Epileptic Disorders
MENUUsing anxiolytics in epilepsy: neurobiological, neuropharmacological and clinical aspects Volume 18, issue 3, September 2016

Despite the increasing interest in psychiatric problems in epilepsy, rather limited literature is available on anxiety disorders (Hamid et al., 2011) and this is probably due to the high comorbidity rates with mood disorders which obscure the distinctive features of anxiety (Jones et al., 2005). However, it seems rather evident that anxiety disorders are frequently present in patients with epilepsy (Brandt et al., 2010), leading to a significant impact on quality of life (Jacoby et al., 2015), and these probably play a key role in suicidality among depressed patients (Placidi et al., 2000).
From an evolutionary perspective, anxiety represents a normal adaptive response to threat or stress, and is characterised by a set of preformed behavioural patterns in response to visual, auditory, olfactory, or somatosensory stimuli (Price, 2003). In addition, anxiety may also be the result of cognitive processes mediating the anticipation, interpretation, or recollection of perceived stressors and threats. From a neurobiological perspective, the Pavlovian-fear conditioning and fear-potentiated startle response, are well-known models of anxiety and they have been extensively used to study anxiety, largely because of their amenability to experimental manipulation (Martin et al., 2009). These neurobiological models have clarified an extended anatomical brain network which centres on the amygdala and a number of connected structures, including mesiotemporal cortical structures, the sensory thalamus and cortices, the orbital and medial prefrontal cortex, the anterior insula, the hypothalamus, and multiple brainstem nuclei (Martin et al., 2009).The amygdala is the determinant in the experience of fear and its autonomic and endocrine response (through the output to the hypothalamus). The output to the periaqueductal grey nuclei is implicated in avoidance behaviour, while the hippocampi play a role in the re-experiencing of fear (Nees and Pohlack, 2014; Cacciaglia et al., 2015). Spontaneous activation of fear circuits is the major hypothesis for explaining symptoms in anxiety disorders and the reduction of such an excessive output from these neurons represents the main target of anti-anxiety treatments. In this regard, it is interesting to note that such a mechanism has a number of similarities with the excessive outburst typical of epileptic neurons, and easily accounts for why some of the agents used in the treatment of epilepsy are also effective against anxiety, and vice versa (Mula et al., 2007).
The term “anxiolytics” refers to a class of compounds that are used to treat anxiety symptoms (Stahl, 2008). They are often discussed together with hypnotics as most of them are also used to induce sleep (table 1). Anxiolytics were traditionally named “minor tranquilizers” to distinguish them from neuroleptics or antipsychotics which were defined as “major tranquilizers”. Traditionally, anxiolytics were thought to interact only with the GABA-A receptor complex but, over time, neuropharmacological studies have shown a number of other neurochemical pathways which can improve anxiety symptoms, either directly or indirectly, through modulation of GABAergic neurotransmission (i.e. voltage-gated calcium channels, alpha andbeta adrenergic receptors, and serotonin neurotransmission). This article is aimed at discussing the neurobiological and neuropharmacological aspects of anxiolytic medications with special attention to epilepsy.
Molecular targets
GABA-A receptors
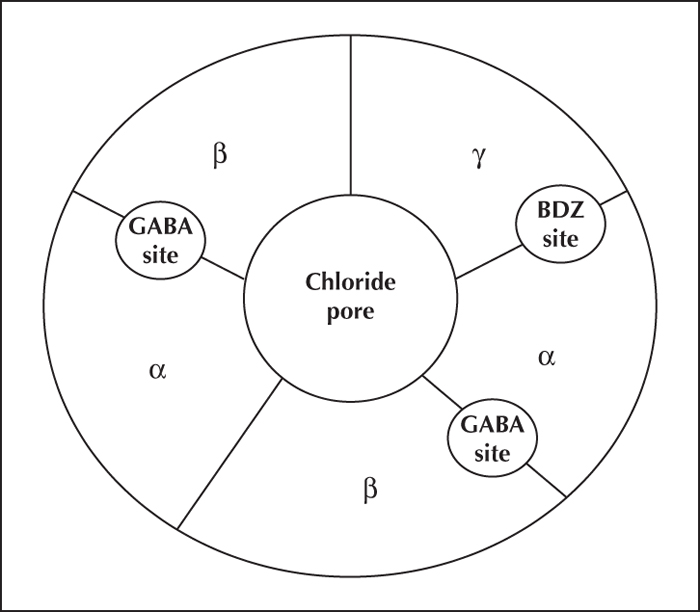
GABA is the principal inhibitory neurotransmitter of the brain and, along with serotonin and noradrenaline, is one of the key targets for anxiolytics (Mula et al., 2007). The GABA-A receptor has a heteropentameric structure surrounding a central pore and is made up of different subunits (Möhler et al., 2001; Mula, 2011) (figure 1). GABA-A receptors can be categorised into three groups based on their alpha isoform content: alpha 1-containing receptors have greatest sensitivity towards BZs (type I); alpha 2, 3 and 5-containing receptors have similar but distinguishable properties (type II); and alpha 4- and 6-containing assemblies have very low BZD affinity (Möhler et al., 2001). GABA-A receptors containing the delta subunit, instead of the gamma, are considered BZD insensitive. In terms of brain localisation, for example, GABA-A receptors containing the alpha 1 subunit are densely represented in the cerebral neocortex (throughout), the hippocampus (DG, CA1 and CA2), and cerebellum, while the alpha 4 and 6 subunits are typical of cerebellar granule cells (Saari et al., 2011). Mutations or genetic variations of the genes encoding the alpha 1, alpha 6, beta 2, beta 3, gamma 2, or delta subunits (GABRA1, GABRA6, GABRB2, GABRB3, GABRG2, and GABRD, respectively) have been associated with a number of epileptic syndromes, from genetic (idiopathic) generalized epilepsies (e.g. juvenile myoclonic epilepsy and childhood absence epilepsy) to severe epileptic encephalopathies, such as Dravet syndrome or Lennox-Gastaut syndrome (Hirose, 2014).
The neurobiology of GABA-A receptors also accounts for why different compounds have a different spectrum of activity. For example, the sedative-hypnotic effect and the partial anti-seizure effect are due to the allosteric positive modulation of GABA-A receptors containing the alpha 1 isoform, while the anxiolytic effect seems to be related to those containing the alpha 2 isoform (Mula, 2011). Non-benzodiazepine (BZD) compounds, such as zolpidem, are often selective for the alpha 1 and alpha 5 isoforms and are, therefore, sedative-hypnotic with partial anti-seizure properties (Mula, 2011). However, it is important to emphasise that all these subunits are subjected to gene variation, and different polymorphisms may be responsible for different anticonvulsant responses to BZDs.
In addition to the biochemical structure of GABA-A receptors, GABA-mediated inhibitory neurotransmission can be classified as “phasic” or “tonic” (Mula, 2011). Phasic inhibition is a short-lasting inhibition, typically generated by the activation of GABA-A receptors containing the gamma subunit and following action potentials in a presynaptic interneuron. Tonic inhibition is represented by GABA-A conductance, activated by ambient GABA in the extracellular space (Farrant and Nusser, 2005), and is mediated by molecularly and functionally specialized GABA-A receptors containing the delta subunit. Tonic inhibition is a long-lasting form of inhibition and does not seem to be affected by the tolerance phenomenon typical of GABA-A receptor stimulation (Farrant and Nusser, 2005; Mula, 2011). The concept of tonic inhibition has received increasing attention in the recent literature. Neurosteroids target GABA-A receptors containing the alpha 4 and alpha 6 isoforms, which tend to co-localize with the delta subunit, and are therefore BZD insensitive (Bianchi et al., 2002). For this reason, neurosteroids have a reduced liability to tolerance and their potential usefulness in both epilepsy and anxiety is receiving increasing attention (Mula, 2011). Interestingly, other well-known antiepileptic drugs (AEDs) seem to increase tonic GABA-ergic neurotransmission, namely phenytoin (Wong and Teo, 1986) and lamotrigine (Wang et al., 2002).
Finally, for completeness, it is worth mentioning GABA-B and GABA-C receptors although they do not seem to play a role in anxiety. GABA-B receptors are expressed presynaptically at GABAergic and glutamatergic synapses and decrease neurotransmitter release by reducing calcium influx. Baclofen is a classic GABA-B receptor agonist because it is specific to GABA-B receptors and does not activate GABA-A receptors. GABA-B specific agonists promote spike-wave discharges, while antagonists (i.e. phaclofen) suppress them, in rodent models of absence epilepsy (Manning et al., 2003).
GABA-C receptors are characterised by the rho subunit and play a unique functional role in retinal signal processing (Zhang et al., 2001).
Noradrenaline and serotonin neurotransmission
Both noradrenaline and serotonin neurotransmission play an important adaptive function in responding to threat or stress.
Noradrenaline increases vigilance, modulates memory, mobilizes energy stores, and elevates cardiovascular function. Nevertheless, these biological responses to threat and stress can become maladaptive if they are chronically or inappropriately activated. Exposure to various types of stressful stimuli increases central noradrenergic (NE) function, especially in the locus coeruleus (LC), the hypothalamus, the hippocampus, the amygdala, and the cerebral cortex(Samuels and Szabadi, 2008). The firing activity of LC neurons also increases during exposure to fear-conditioned stimuli and other stressors or threats (Steckler et al., 2005). The recurrent symptoms of anxiety disorders, such as panic attacks, insomnia, exaggerated startle, and chronic sympathetic autonomic arousal, may conceivably reflect elevated NE function. Patients with post-traumatic stress disorder (PTSD) and panic disorder (PD) show evidence of heightened peripheral sympathetic nervous system arousal which, because of the correlation between peripheral sympathetic activity and central noradrenergic function, is compatible with the hypothesis of increased central NE activity in these disorders (Blechert et al., 2007). BZDs decrease LC neuronal firing activity but other agents, which specifically target noradrenergic receptors, such as beta or alpha blockers, are successfully used in the treatment of anxiety symptoms.
On the contrary, the role of serotonin (5-HT) neurotransmission in anxiety disorders is still a matter of debate, and normal and altered 5-HT receptors and/or serotonin transporter (SERT) function have been demonstrated in an equal number of studies (Maron et al., 2004a, 2004b; Freitas-Ferrari et al., 2010). During exposure to fear-conditioned stimuli, the 5-HT turnover in the medial prefrontal cortex correlates with the severity of stress and stimulates both anxiogenic and anxiolytic pathways within the forebrain, depending on the region involved and the 5-HT receptor subtype that is predominantly stimulated. A well-known and influential hypothesis regarding the involvement of the serotoninergic system in anxiety postulates that 5-TH2A receptors of the amygdala mediate the anxiogenic effects, while 5-HT1A receptors in the hippocampi provide resilience to aversive stimuli (Graeff et al., 1993). This is confirmed by the 5-HT1A receptor knock-out animal model which exhibits anxiety behaviour and the anxiolytic effect of 5-HT1A receptor agonists (Ramboz et al., 1998). In this regard, it is important to note that glucocorticoids modulate the genetic expression of both 5-HT1A and 5-HT2A receptors (Watanabe et al., 1993; López et al., 1998). In fact, gene expression of post-synaptic 5-HT1A receptors in the hippocampi is down-regulated by corticosteroids, whereas 5-HT2A receptors seem to be up-regulated (Watanabe et al., 1993; López et al., 1998). This mechanism would explain the number of plastic changes and brain network abnormalities in patients with anxiety disorders and the role of acute and chronic stress associated with such changes. Still, this would also explain why selective serotonin re-uptake inhibitors (SSRIs), or other serotoninergic antidepressants, are successful for the long-term treatment of anxiety disorders rather than being anxiolytics per se.
Voltage-gated ion channels
Voltage-gated ion channels have always been popular in the psychiatric literature (Gargus, 2006). The different types, primarily recognised as Na+, K+, Ca2+, Cl-, have been basically associated with neuronal firing and processes. Therefore, any drug targeting ion channels can influence all systems related to neuronal activity. In the context of anxiety, the calcium channels involved are those that have received increasing attention, as they have been shown to improve anxiety symptoms in animal models of anxiety (Mula et al., 2007; Zamponi, 2016).
Calcium channels consist of two families: high voltage-activated (HVA) and low voltage-activated (LVA). The HVA family comprises L-type (generating a long-lasting current) and N-, P-, Q-, and R-type channels (expressed in nerve terminals and responsible for the calcium entry that triggers neurotransmitter release) (Mula, 2009). They are heterotrimeric structures consisting of three subunits: alpha, beta, and alpha 2-delta. The alpha subunit forms a pore with the ancillary subunit beta, while the alpha 2-delta subunit forms a functional pore by linking with the subunit alpha. The LVA family consists only of T-type channels which are monomers and composed of only the alpha subunit.
LVA and HVA differ in function, localisation, and electrophysiological activity. Although this represents an oversimplification, LVA T-type channels generate transient currents, have a somatodendritic localization, and are critical to pacemaker activity and some patterns of burst firing, while HVA channels are more likely to be implicated in neurotransmitter release. There is some evidence that calcium channels, particularly HVA channels, may be implicated in the pathophysiology of mood disorders (Lodge and Li, 2008).Genetic variation in CACNA1C, a gene encoding the alpha 1C subunit of the L-type voltage-gated calcium channel, has been associated with bipolar disorder, depression, and schizophrenia (Bhat et al., 2012). It is also well known that calcium channel modulators can have either depressogenic or antidepressant properties (Perucca and Mula, 2013). Drugs targeting the alpha 2 delta subunit, such as gabapentin or pregabalin, seem to have anxiolytic properties (Joshi and Taylor, 2006), but the reason for this still remains unexplained. One possibility is that the blockade of HVA calcium channels translates into a reduction in glutamate release (Farber et al., 2002), which may be ultimately responsible for positive effects on mood and some antianxiety properties. However, further studies are needed. At any rate, there is enough clinical evidence (see below) to support their use in anxiety disorders.
Specific anxiolytic agents
Benzodiazepines
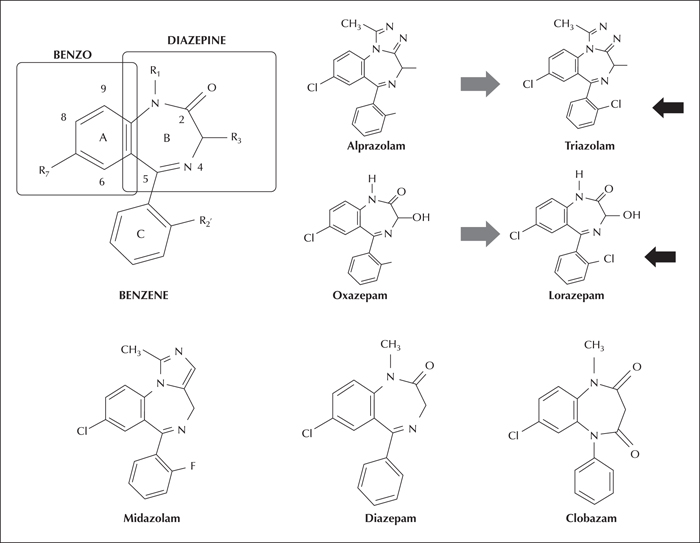
BZDs are a class of drugs chemically characterised by a benzene and a diazepine ring fused together, plus a third benzene ring (figure 2). Chlordiazepoxide was the first synthetized BZD and was accidently discovered by Leo Sternbach in 1955. It was also the first BZD introduced into clinical practice and made available by La-Roche in 1960. Afterwards, numerous different BZDs were synthetized and approximately 30 of them are currently available in clinical practice as important compounds, not only as anxiolytics or hypnotics but, most importantly, for the treatment of status epilepticus, epileptic seizures, and, in general, anaesthesia (Saari et al., 2011).
Clinicians are often unaware of the wide range of BZDs available on the market and how they differ in terms of activity and clinical effects (Stahl, 2008; Schatzberg and Nemeroff, 2009). BZDs can be divided into a number of subgroups according to different chemical and pharmacological parameters (figure 2).
The majority of compounds which are well known to clinicians belong to the 1,4 group, while clobazam is the only 1,5 BZD. The difference in the chemical structure between diazapam (1,4 BZD) and clobazam (1,5 BZD) is shown in figure 2. The main difference between 1,4 and 1,5 BZDs is in the hypnotic effect (Nicholson, 1979). In fact, while all 1,4 BZDs have more or less a significant hypnotic effect, 1,5 BZDs lack such an effect, which accounts for why clobazam has a different impact on cognitive functions, compared to diazepam (Bawden et al., 1999). As a note, both clozapine and olanzapine are also 1,5 BZDs but their chemical structure is, however, completely different from the group of compounds referred to as “BZDs”. Another important BZD subgroup is that of imidazo-BZDs; for example, midazolam (figure 2). Their distinctive feature is mainly based on a chemical point of view, because they exhibit pH-dependent water solubility. In fact, below pH 4, they are freely water-soluble, while at physiological pH in plasma, the ring closes and the drug becomes lipid-soluble and rapidly penetrates the blood/brain barrier to exert their action. They are, therefore, more water-soluble/stable than other BZDs.
Finally, BZDs can also be classified as short-, intermediate- or slow-acting, according to pharmacokinetic and pharmacodynamic parameters (table 2) (Stahl, 2008). Short-acting BZDs are hypnotics and can be used to induce general anaesthesia, while slow-acting BZDs are recommended for the treatment of anxiety.In this context, it should be acknowledged that some BZDs undergo complex and extensive metabolism, leading to a number of metabolites which, in some cases, are active and contribute to the final pharmacological effect. For example, BZDs, such as flurazepam, present a mixed profile; they are short-acting in terms of a rapid hypnotic effect, but the pharmacological activity is relatively sustained due to the long half-life of the active metabolite (table 2). An extensive discussion on the pharmacokinetics of BZDs is beyond the aim of this article, however, the reader should bear in mind that individual differences in the metabolism of BZDs may be responsible for inter-individual differences in the magnitude of the effect and the onset of side effects. In fact, the elimination half-life of diazepam, like other long-half-life BZDs, is twice as long in the elderly compared to younger individuals, and doctors should always adjust the dosage according to age.
Antiepileptic drugs
During the last 15 years, clinical researchers have become increasingly interested in the potential for AEDs to improve or control anxiety symptoms. This was due to the limitations connected with the long-term use of BZDs (see below) and the number of patients still refractory to first-line treatment. In addition, as already briefly discussed, the spontaneous activation of fear circuits has a number of commonalities with the spontaneous activation of brain networks described in epilepsy. For all these reasons, a number of AEDs have been trialled for anxiety disorders (Mula et al., 2007). Despite a considerable number of published studies, the majority have several methodological limitations: inadequate sample size, lack of a placebo control, use of non-specific outcome measures (i.e. the clinical global impression scales), and lack of control for concomitant bias (i.e. comorbidities, diagnostic subtypes, and concomitant medications). These factors may help explain why AEDs have yielded inconsistent results in the treatment of anxiety disorders. At present, the most convincing data to support the use of an AED to improve or control anxiety symptoms is for pregabalin (at dosages between 300 mg and 600 mg) in patients with generalized anxiety disorder, with or without comorbid depression (Feltner et al., 2003; Pande et al., 2003, 2004; Montgomery et al., 2008; Diaper et al., 2013). Pregabalin (Pande et al., 2004; Kawalec et al., 2015) and gabapentin (Pande et al., 1999) have also shown promising results for the treatment of social phobia, but further studies are needed. For the remaining AEDs, results are still preliminary and randomised controlled trials are lacking (Mula et al., 2007).
Antidepressants
Antidepressants are increasingly used for the treatment of anxiety disorders (Bandelow et al., 2008) and this is due to the number of limitations associated with BZDs, such as tolerance, dependence, and the risk of withdrawal (see below). Data is available mainly for SSRIs and serotonin and noradrenaline re-uptake inhibitors (SNRIs) for long-term treatment. It remains controversial why antidepressants are effective against anxiety, but this seems to be related to the number of plastic changes to noradrenergic and serotoninergic neurotransmission. For panic attack disorder, SSRIs were shown to be as effective as tricyclics, but better tolerated (Bakker et al., 2002). Data on generalised anxiety disorder has focused mainly on venlafaxine, paroxetine, and imipramine, and all of these have shown good efficacy, but overall appear to be as effective as lorazepam or pregabalin (Mula and Strigaro, 2010). For social anxiety disorder and post-traumatic stress disorder, data are still limited and further studies are needed, although there is promising evidence for sertraline and paroxetine (Mula and Strigaro, 2010).
Evidence from clinical studies
Anxiety disorders are chronic conditions with a clear relapsing/remitting course, and this has been demonstrated by a number of cross-sectional and prospective studies (Maser, 1990). However, this concept is of great relevance as clinicians remain focused almost entirely on the acute control of anxiety symptoms and only secondarily acknowledge relapse prevention. In addition, the natural history of anxiety disorders is frequently complicated by Axis I (e.g. major depression, bipolar disorder, psychoses, etc.) and Axis II (i.e. personality disorders) comorbidities which have a major impact on response to treatment. For example, 73% of patients with panic attacks have other comorbid conditions, ranging from major depression to substance abuse and personality disorders (Maser, 1990) that need to be taken into account in any long-term anxiolytic treatment. BZDs have been historically considered first-line treatment for the acute management of anxiety, but their long-term use should be avoided as BZDs may lead to complications, such as abuse liability, dependence, and withdrawal syndrome. Long-term use is usually defined by a period of daily use over at least three months (Voshaar et al., 2006), because this seems to be long enough to cause changes in neural adaptation that counteract the drug's effects (tolerance phenomenon). In these subjects, even a reduction in dose may cause rebound symptoms that are almost identical to those for which the drug was initially taken (e.g. insomnia, agitation, and panic attacks). In epilepsy, tolerance and dependence are even more relevant as seizures are common manifestations of a withdrawal reaction. In general terms, long half-life BZDs should be preferred relative to short-acting compounds, due to less associated rebound symptoms, but the management of withdrawal should be planned on a case by case basis, depending on age, concomitant comorbidities, and seizure risk. The potential risk of dependence with pregabalin should be considered, however, compared to BZDs, this is less evident (Caster et al., 2011).
Data on treatment of anxiety disorders in epilepsy are still limited and rely heavily on clinical experience (Kerr et al., 2011). However, as already mentioned, it is evident that anxiety disorders represent a frequent comorbidity in patients with epilepsy (Brandt et al., 2010). The Commission on Neuropsychiatry of the International League Against Epilepsy published a collection of articles concerning treatment strategies in adults with epilepsy and psychiatric disorders (Mula and Kanner, 2013), one of which was dedicated to the treatment of anxiety disorders (Mula, 2013a). Evidenced-based therapeutic strategies in patients with anxiety disorders without epilepsy can be easily adapted to patients with epilepsy by considering specific needs (table 3). For panic disorder, a combined approach, namely SSRIs and cognitive behavioural therapy (CBT), is recommended for the acute phase, while long-term maintenance treatment can be combined or based on CBT alone, depending on the individual patient. For generalized anxiety disorder (GAD), pregabalin can be reasonably considered first choice for the acute and long-term maintenance treatment as it is licensed, although not everywhere, for both conditions. For social anxiety disorder and post-traumatic stress disorder, SSRIs, in particular sertraline and paroxetine, should be preferred for their low risk of interactions and favourable tolerability. For obsessive compulsive disorder (OCD), CBT should always be considered as first-line treatment. When drug treatment is needed, SSRIs, in particular sertraline at 100 mg, is preferred. Although it is reasonable to embrace standardized treatment protocols that have been developed for people with anxiety without epilepsy, it is also evident that psychiatric disorders associated with epilepsy present, more often than not, with atypical features (Mula, 2013b) that may require individualised approaches. For this reason, studies on patients with epilepsy are urgently needed.
Regarding children with epilepsy, data are even more limited, relative to that of adults. This is fairly surprising if we consider that, in the general population, anxiety disorders are much more common in children than in adults (Costello et al., 2005; Franz et al., 2013). In addition, children with anxiety disorders seem to be at an increased risk of further psychiatric comorbidities, such as ADHD or conduct disorder (Kendall et al., 2010). Finally, it seems now established that half of adults with anxiety or depression have a history of anxiety onset before the age of 15 (Kim-Cohen et al., 2003). For all these reasons, The American Academy of Child and Adolescent Psychiatry has recommended that children and adolescents are routinely screened for symptoms of anxiety (Connolly et al., 2007). It would therefore appear evident that both careful assessment and prompt treatment for children with epilepsy would probably reduce the development of major problems during adulthood. Jones (2014) recently reviewed the management and treatment of anxiety disorders in children and adolescents with epilepsy. SSRIs remain the first-line treatment, especially for OCD, and there is promising data for venlafaxine for the treatment of GAD and separation anxiety disorder. Again, PGB represents a good therapeutic option, but there are no studies on PGB in children with epilepsy and anxiety disorders.
Finally, it is important to mention that children may present paradoxical reactions to short-acting BZDs (Jackson et al., 2015). This is a well-known phenomenon; for example, midazolam when used to induce mild sedation for elective surgical/invasive procedures (McKenzie and Rosenberg, 2010). Older patients with intellectual disabilities are affected similarly to children and may develop paradoxical agitation and aggressive behaviour with short-acting BZDs (Barron and Sandman, 1985). For this reason, short-acting BZDs should be avoided or carefully used in these patients.
Conclusions
Anxiolytic treatment is often perceived by clinicians to be safe and easy. However, a multitude of different compounds with different mechanisms of action and peculiarities are available.In the context of epilepsy, this is even more relevant, as the main target for most of these medications is exactly the same as that for AEDs. Controlled studies in both children and adults with epilepsy are urgently needed in order to develop tailored treatment strategies for anxiety disorders in this specific subgroup of patients.
Acknowledgements and disclosures
The author has not received any financial support for the present article, but has previously received consultancy fees from UCB Pharma, Eisai, Pfizer and Elsevier, as well as support from Bial and Special Products Ltd.