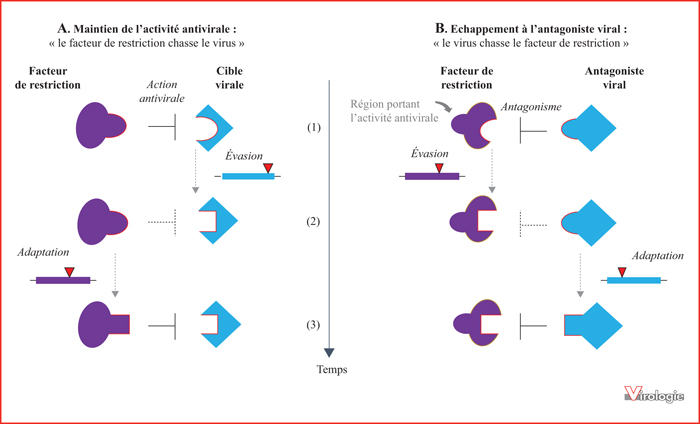
Conflit génétique entre les protéines antivirales et le virus. Les protéines de l’hôte et les protéines virales sont engagées dans une course évolutive rapide qui suit l’hypothèse de la Reine rouge [9]. Au cours de l’évolution, les deux entités subissent une série d’adaptations et de contre-adaptations, où la sélection de mutations adaptatives d’une entité va exercer une pression de sélection diversifiante sur son adversaire, qui évoluera alors à son tour. A) Le facteur de restriction est ici en position « d’attaque ». Il est capable de reconnaître une cible virale et de bloquer la réplication du virus. Seuls les gènes antiviraux capables de reconnaître la cible virale seront sélectionnés au cours du temps. Dans ce scénario, le virus est en position d’échappement. B) Le facteur de restriction est ici en position « défensive » face à un antagoniste viral. Seuls les facteurs de restriction qui échappent à la reconnaissance de l’antagoniste viral seront sélectionnés au cours du temps. L’interface virus-hôte est représentée par une ligne rouge. Les changements de forme à l’interface représentent des changements de la structure protéique. Ces changements en acides aminés sont le résultat de mutations non synonymes représentées par des triangles rouges sur le schéma du gène correspondant. Basé sur des concepts présentés dans [7, 8].
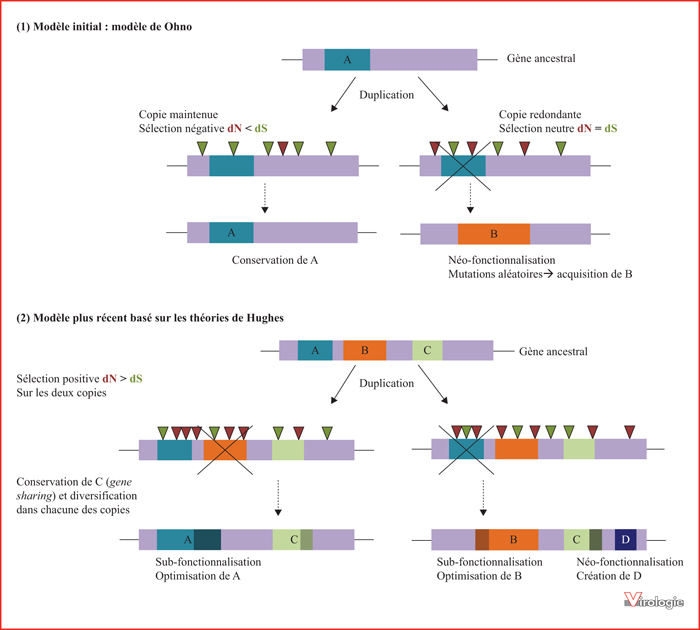
Duplication, sélection et innovations génétiques selon le modèle initial de Ohno (1) et celui basé sur les théories de Hughes (2). (1) Exemple du modèle initial de Ohno [12, 13]. Le gène ancestral code pour une fonction A. Suite à la duplication et pendant la phase de divergence des copies, la copie redondante est soumise à une pression de sélection neutre (dN = dS), alors que la copie maintenue est sous pression de sélection purifiante (dN < dS). L’accumulation de mutations aléatoires entraîne le plus souvent la pseudogénisation de la copie redondante. Cependant, dans certains cas, ces mutations entraînent la néo-fonctionnalisation de la nouvelle copie, représentée ici par l’acquisition de la fonction B. (2) Exemple du modèle basé sur les théories de Hughes [14, 15]. Le gène ancestral est multifonctionnel ou code pour plusieurs domaines d’interaction avec des protéines endogènes ou des protéines virales. Après la duplication, les deux copies sont soumises à une pression de sélection diversifiante (dN > dS). Les copies se spécialisent et chacune optimise, par exemple, une des régions ancestrales, A ou B. La région C est conservée chez les deux copies (possible « effet dose » [14]) et/ou sa fonction est diversifiée dans chacune des copies. L’accumulation de mutations non synonymes entraîne l’émergence de nouvelles fonctions, D sur l’une des copies. Les zones représentées plus foncées symbolisent la spécialisation ou la divergence des régions par rapport aux régions ancestrales. Les triangles verts symbolisent des mutations synonymes ; les triangles rouges des mutations non synonymes ; A, B, C et D peuvent coder des fonctions cellulaires, des fonctions antivirales ou encore des interfaces protéiques en contact avec des protéines virales.
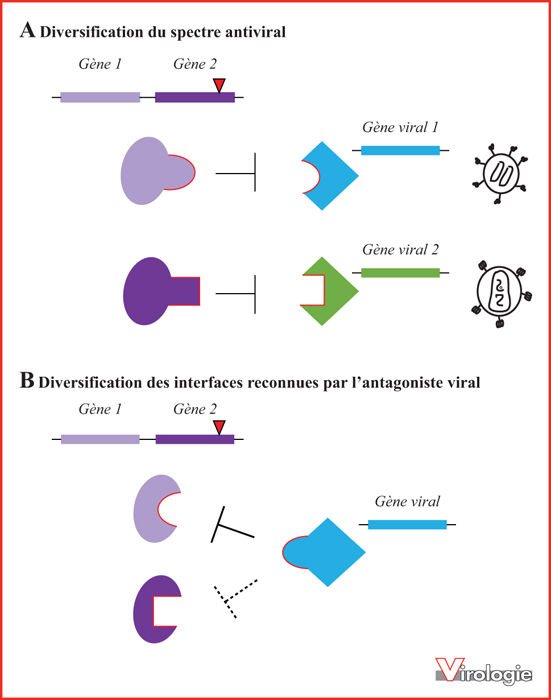
Modèle des bénéfices potentiels apportés par la duplication des « gènes antiviraux ». A) La conservation des deux paralogues a été sélectionnée car la divergence des deux gènes a entraîné l’acquisition de nouvelles activités antivirales. Ainsi, chaque protéine antivirale codée peut interagir avec une protéine cible virale différente, provenant potentiellement de différents virus, élargissant ainsi le spectre antiviral. B) La conservation des deux paralogues a été sélectionnée car la divergence des deux copies de gènes a permis la diversification des interfaces reconnues par l’antagoniste viral (i.e., le virus est contraint dans son évolution).
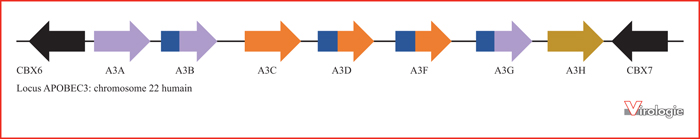
Le locus APOBEC3 chez l’homme a une histoire évolutive complexe incluant plusieurs duplications, fusions, et divergences de gènes. Chez l’homme, les sept gènes paralogues APOBEC3s sont flanqués de part et d’autre par les gènes CBX6 et CBX7 (en noir) sur le bras long du chromosome 22. En violet, les gènes ou fragments de gènes provenant du gène ancestral A3Z1 ; en bleu et orange, les gènes ou fragments de gènes provenant du gène ancestral A3Z2 ; en ocre, le gène provenant du gène ancestral A3Z3[22]. La plupart des mammifères présentent une organisation similaire du locus.
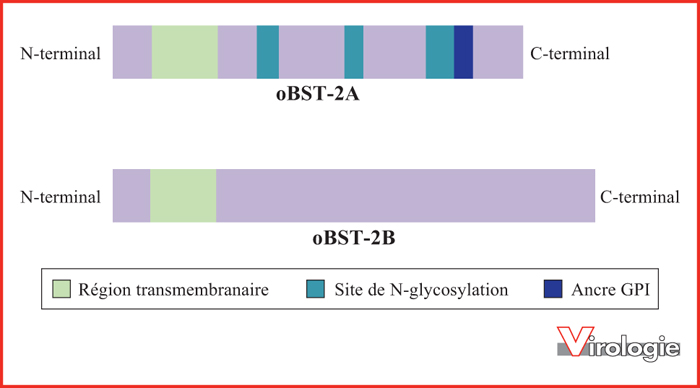
Les protéines oBST-2A et oBST-2B des ovins ont des caractéristiques structurales et des domaines de modifications post-traductionnelles distincts. Les deux protéines ovines issues de deux gènes paralogues possèdent une région transmembranaire en N-terminal (en vert). oBST-2B ne présente pas les sites de N-glycosylation et l’ancre GPI retrouvés, respectivement, sur la portion extracellulaire et en C-terminal sur oBST-2A.
The antiviral proteins, also known as restriction factors, are the primary cellular defense against viral pathogens. These proteins from the innate immune system are in direct interactions with viral proteins. These antagonistic interactions come in two flavors: restriction factors are able to directly target the virus to restrict its replication and/or they may be the target of viral antagonists. Such long-term antagonistic virus-host interactions have set up an evolutionary “arms-race” between the two adversarial entities. This genetic conflict leads to a rapid evolution of viruses and antiviral genes. In particular, amino acids at the virus-host interface change more frequently than expected over time. On the other hand, more drastic host genome modifications have also been selected over time to counteract the rapid evolution of viruses. Reflecting the fact that numerous restriction factors belong to a gene family, it appears that duplication of antiviral genes has occurred frequently during the course of evolution. Here, we will review how duplication of antiviral genes has been selected in the host and how the divergence and plasticity of the duplicated genes may have been advantageous in the virus-host genetic conflict. We will also briefly expose the limits to such innate immune gene expansion.