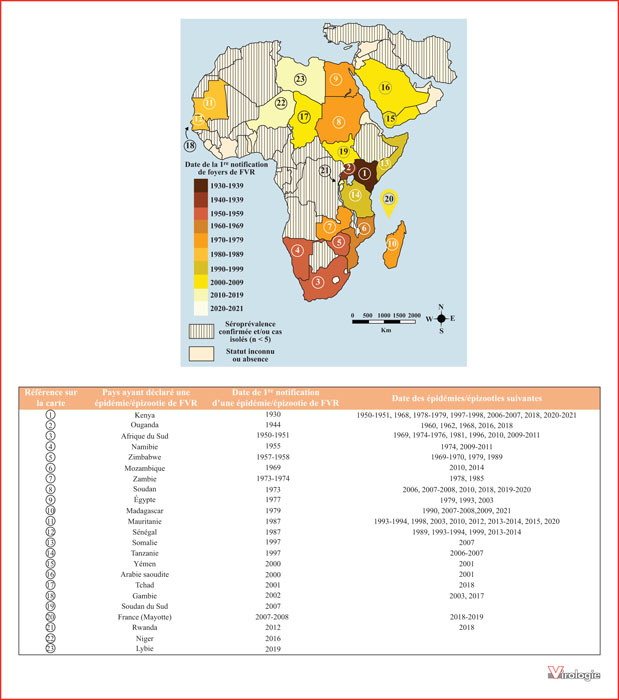
Distribution spatio-temporelle de la FVR entre 1930 et 2021. Le dégradé de couleurs représente les décennies durant lesquelles a été notifiée la première épidémie et/ou épizootie liée à la FVR sur un territoire donné. Les pays hachurés correspondent aux pays sans épidémie/épizootie mais où la circulation virale a été décrite directement ou indirectement par des études sérologiques. Le tableau présente la liste des épidémies/épizooties déclarées en Afrique et au Moyen-Orient [7, 8, 15, 17, 19-22].
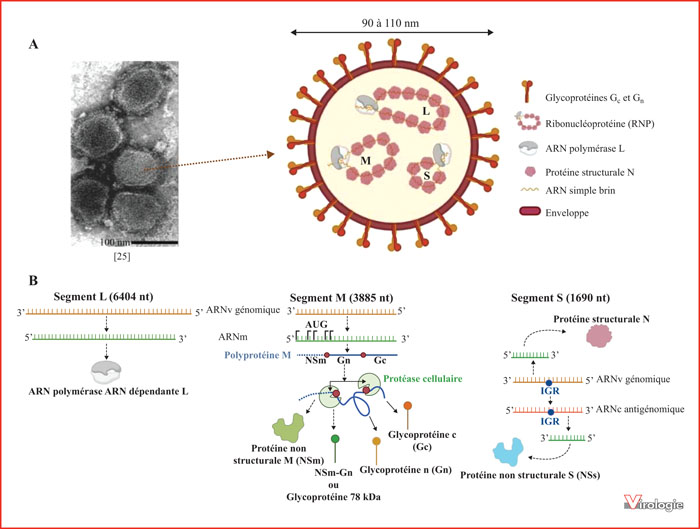
A.Particules virales du RVFV obtenues par microscopie électronique à transmission [25] et son organisation schématique. B. Représentation schématique de la stratégie d’expression du génome viral. Les ARNs viraux de polarité négative présents dans la particule virale vont être exprimés différemment en fonction du segment. Le segment L va être transcrit en ARNm (en vert) puis traduit pour permettre la synthèse de la polymérase L. Le segment M va également être transcrit en ARNm, qui possède plusieurs codons start (AUG). Par un mécanisme de leaky-scanning (saut de codon start), la traduction de cet ARNm va permettre la formation de la polyprotéine M de taille variable (variabilité représentée par les pointillés bleus). La présence de sites de clivage (point rouge) sur cette polyprotéine permet, par l’action de protéases cellulaires, de la cliver en plusieurs protéines. Ainsi, en fonction de la protéine M synthétisée et des sites de clivage, il peut y avoir synthèse des protéines de surface Gc et Gn, et soit la glycoprotéine 78 kDa, soit la protéine non structurale M. Le segment S est ambisens : le brin génomique va permettre la formation d’un ARNm permettant la synthèse de la protéine structurale N et le brin antigénomique va permettre la formation lui aussi d’un autre ARNm, qui permettra la synthèse de la protéine non structurale S. La transcription de ces ARNm est régulée par la présence de la région IGR (intergenic region, point bleu) qui permet la terminaison de la transcription.
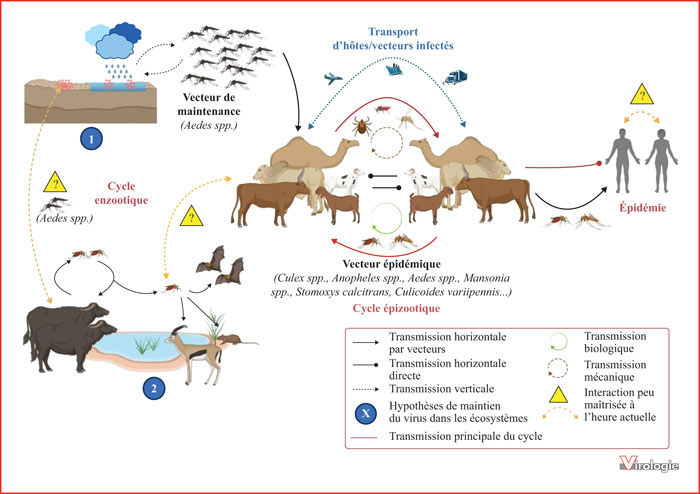
Représentation schématique du cycle de transmission biologique du RVFV. Au sein du cycle enzootique, il existe deux hypothèses du maintien du virus dans l’environnement entre les épidémies : (1) par transmission verticale puis dans l’environnement par oviposition chez certaines espèces de moustiques du genre Aedes (vecteur de maintenance), et/ou (2) par l’existence d’un cycle sylvatique entre une ou plusieurs espèces animales réservoirs et certaines espèces de moustiques vectrices. La source d’infection des élevages de ruminants est d’origine environnementale (prolifération des vecteurs Aedes) ou anthropique (mouvements de populations…). La transmission du virus au sein des élevages par des vecteurs arthropodes épidémiques permet son amplification puis l’établissement du cycle épizootique. La transmission du virus à la population humaine à partir d’élevages infectés (contact, aliments crus, vecteurs) déclenche une épidémie.
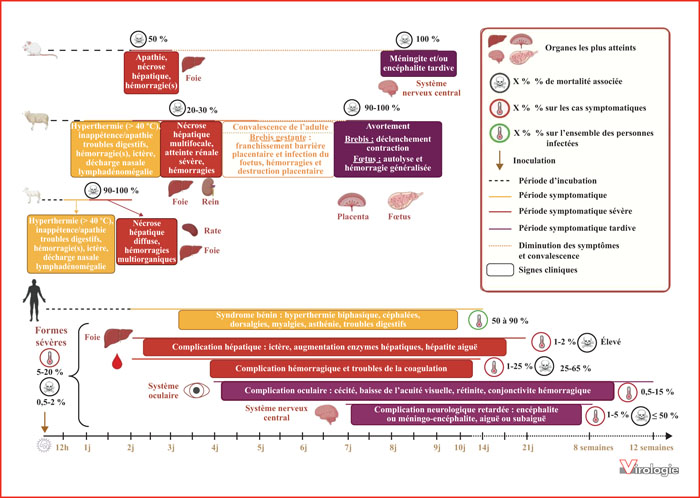
Représentation schématique de la cinétique et de la symptomatologie associées à l’infection par le RVFV en fonction de l’espèce : modèle souris, mouton, agneau, Homme (de haut en bas).
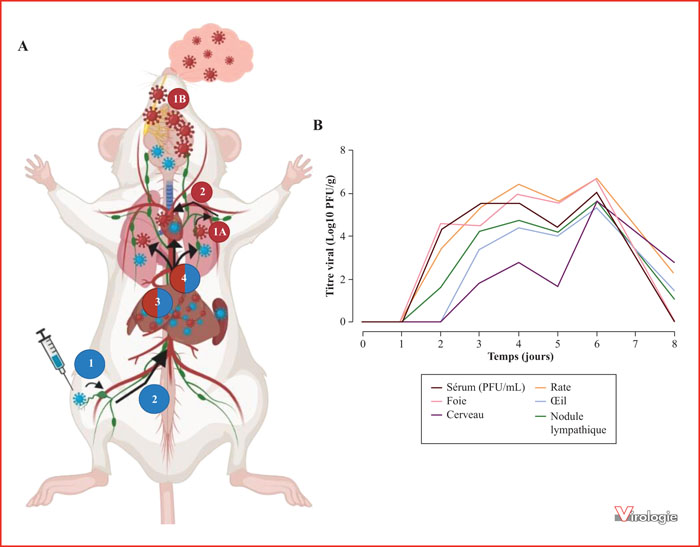
A. Cinétique d’infection du RVFV dans le modèle murininoculé par voie intradermique (bleu), mimant ainsi l’infection par piqûre du vecteur moustique, ou par aérosol (rouge), mimant la transmission directe. Phase précoce : par voie intradermique, le RVFV infecte les cellules de l’immunité naturelle de la peau (cellules dendritiques, macrophages, …) (1) et par aérosols, il infecte les cellules de l’immunité naturelle résidentes du tissu pulmonaire (1A) mais atteint également le système nerveux en infectant le nerf olfactif (1B). Indépendamment de la voie d’exposition, il est ensuite acheminé vers les ganglions lymphatiques qui serviront de site de réplication précoce. Le virus rejoint la circulation sanguine pour atteindre les organes cibles dont le foie (2). Phase hépatique : le foie est le site majeur de réplication, responsable de la production intense de virions, indépendamment de la voie d’exposition (3). Phase tardive : la réplication du RVFV au niveau hépatique augmente la dissémination par voie hématogène du virus vers les tissus secondaires (cerveau, œil, cœur), majorant alors la charge virale et la réplication du virus dans ces tissus (4).
PFU, particle forming unit. (Réalisé à partir des données de [70, 94, 98].)
B. Évolution de la charge virale dans les tissus au cours du temps après infection par le RVFV dans le modèle murin. Dans les deux premiers jours post-infection, une colonisation du compartiment sanguin, des organes lymphoïdes et du foie est observée. Elle est ensuite suivie d’une colonisation plus tardive (48-72 h post-infection) dans d’autres organes tels que l’œil et le cerveau avec un pic de réplication au sixième jour post-infection. À partir du huitième jour post-infection, la charge virale détectable dans les organes colonisés tardivement reste plus élevée que celle détectée en phase précoce dans la rate, le foie et le compartiment sanguin jusqu’à une disparition totale du virus dans ces derniers.
(Réalisé à partir des données de [94].)
La fièvre de la vallée du Rift (FVR) est une arbovirose émergente majeure possédant un cycle de transmission complexe. Le virus de la FVR (RVFV) est transmis aux élevages de ruminants par des moustiques vecteurs, provoquant des épizooties, puis de l’animal à l’Homme, déclenchant alors des épidémies. Au cours de son cycle, le RVFV infecte une large gamme d’hôtes, la pathogenèse associée soulevant de nombreuses questions. Malgré un tropisme principalement hépatique, le RVFV a également un tropisme multicellulaire induisant un effet physiopathologique dans de nombreux tissus. Un tableau clinique commun est habituellement retrouvé (hépatite sévère, hémorragies, leucopénie) mais une variabilité interspécifique a également été décrite. Ainsi, certaines formes sont principalement retrouvées chez l’Homme (atteintes neurologiques et oculaires) ou dans les élevages de ruminants (vagues d’avortements). Bien que les mécanismes moléculaires impliqués soient encore mal connus, il semblerait que la sévérité de la pathologie soit liée à la réponse inflammatoire précoce. De même, la pathogenèse du RVFV semble être soumise à de nombreux facteurs responsables d’une variabilité inter-individuelle importante dans la sévérité de l’infection. Devant l’absence de traitement spécifique, il semble primordial de mieux comprendre la pathogenèse du RVFV.