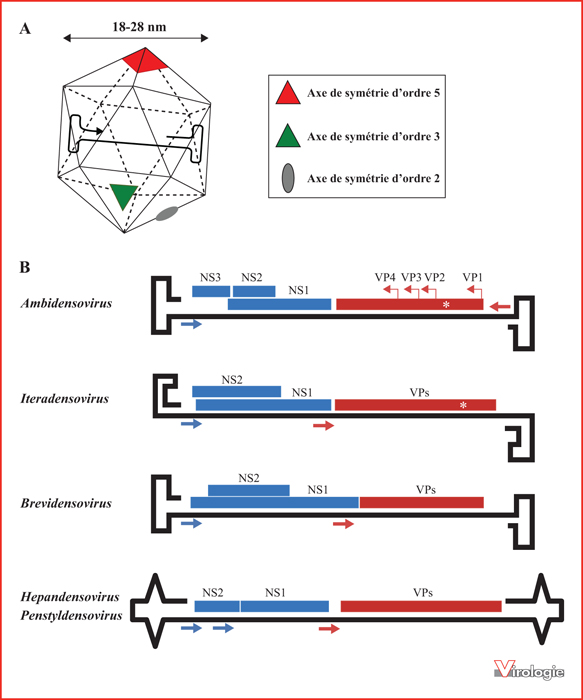
Structure et organisation génomique des Densovirinæ. A) Les densovirus sont des virus nus qui possèdent une capside icosaédrique de 18 à 28 nm de diamètre. Cette capside est constituée par l’assemblage de 60 protéines de structure (VP) et protège le génome viral constitué d’une molécule d’ADN monocaténaire et linéaire [15]. B) Le génome des densovirus est une molécule d’ADN simple brin de 3,7 à 6,3 kilobases, dont les extrémités contiennent des répétitions terminales inversées qui se replient en structures de type « épingles à cheveux » et servent à la réplication virale. Il possède des gènes codant deux types de protéines : les protéines non structurales (NS, en bleu) et les protéines de capside (VP, en rouge). Les flèches bleues et rouges indiquent les promoteurs initiant la transcription des gènes codant, respectivement, les protéines NS et les protéines VP. La localisation du motif phospholipase A2 (PLA2) est indiquée par une étoile. Les virus du genre Ambidensovirus possèdent un génome ambisens, celui des 4 autres genres (Iteradensovirus, Brevidensovirus, Hepandensovirus et Penstyldensovirus) est monosens.
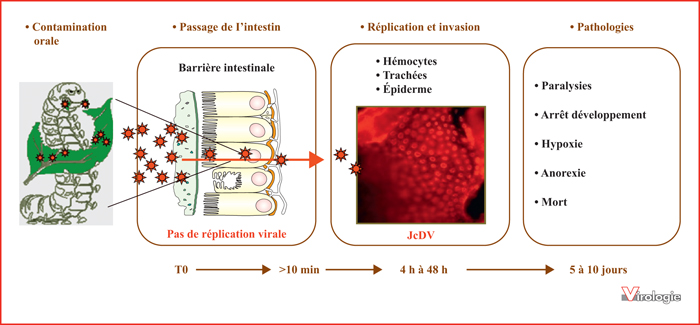
Pathogenèse du densovirus JcDV chez le lépidoptère Spodoptera frugiperda. Les particules virales, ingérées par la chenille via l’alimentation, rejoignent rapidement l’intestin moyen et traverse l’épithélium intestinal sans réplication virale pour atteindre les tissus cibles sous-jacents, principalement les hémocytes, les trachées et l’épiderme. La multiplication virale dans ces tissus bloque les mues et entraîne l’asphyxie des chenilles par obstruction des trachées, ce qui conduit à la mort de l’hôte, 5 à 10 jours après l’ingestion initiale. L’image d’immunofluorescence présentée au centre montre la multiplication virale dans l’épiderme d’une chenille à 5 jours p.i. (tirée de [31]). JcDV (en rouge) est mis en évidence à l’aide d’un anticorps primaire de souris dirigé contre les protéines de capside (VP) et d’un anticorps secondaire anti-souris Alexa Fluor® 594 (Invitrogen).
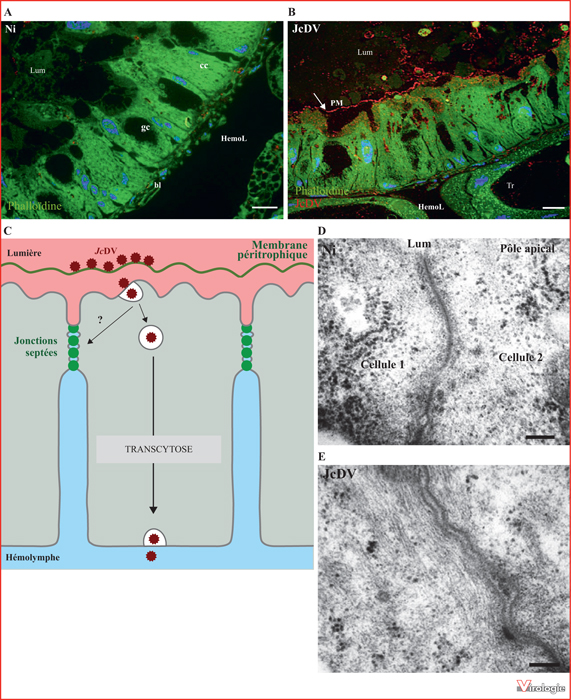
Franchissement de la barrière intestinale du lépidoptère Spodoptera frugiperda par le densovirus JcDV (d’après [49]). A) Image d’immunofluorescence sur coupe semi-fine d’intestin moyen de S. frugiperda montrant l’organisation de l’épithélium intestinal. L’actine (en vert) est visualisée à l’aide de phalloïdine-FITC et les noyaux (en bleu) à l’aide de Hoechst. B) Image d’immunofluorescence sur coupe semi-fine d’intestin moyen de S. frugiperda montrant la localisation des particules virales (en rouge) 15 min après leur ingestion par la chenille. La flèche indique la position de la membrane péritrophique (PM). C) Schéma résumant les mécanismes de franchissement de la barrière intestinale par JcDV. Dans l’intestin moyen, les particules virales se concentrent majoritairement sur la membrane péritrophique et les microvillosités apicales de l’épithélium intestinal. JcDV entre ensuite dans les cellules columnaires par un mécanisme d’endocytose dépendante de la dynamine et de la cavéoline et traverse l’épithélium intestinal par transcytose sans réplication virale. La transcytose du virus entraîne une augmentation de la perméabilité intestinale résultant de l’ouverture partielle des jonctions septées. Les mécanismes précis (directs ou indirects) permettant cette ouverture ne sont pas encore connus. D) Micrographie électronique sur coupe ultrafine d’intestin moyen de S. frugiperda montrant la structure en échelle des jonctions septées reliant les cellules intestinales. E) Micrographie électronique sur coupe ultra-fine d’intestin moyen de S. frugiperda montrant l’ouverture des jonctions septées consécutive à l’infection virale (10 min p.i.).
Barre d’échelle A et B : 20 μm. Barre d’échelle D et E : 100 nm.
Ni : non infecté ; cc : cellule columnaire ; gc : cellule caliciforme ; Lum : lumière de l’intestin ; HemoL : hémolymphe ; bl : lame basale ; Tr : trachée.
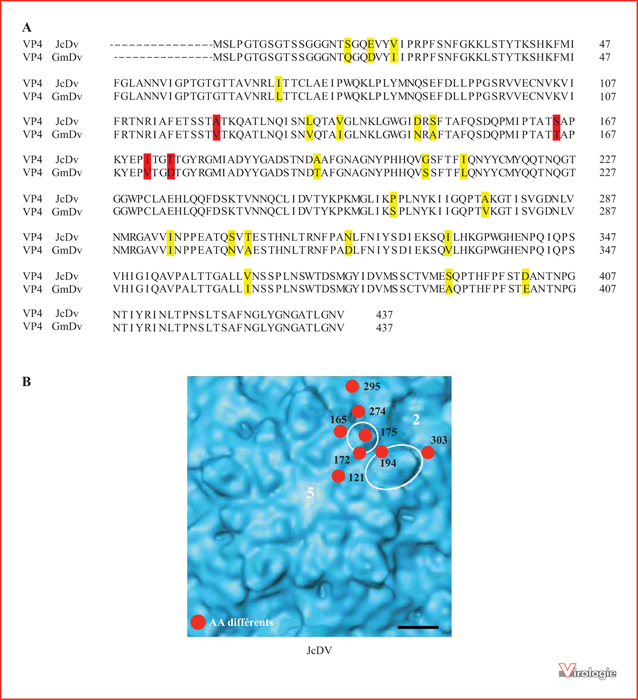
Localisation des acides aminés de la protéine de capside VP4 impliqués dans le tropisme intestinal de JcDV. A) Alignement des séquences protéiques des VP4 de JcDV et de GmDV (437 acides aminés, 94 % d’identité). Les acides aminés qui diffèrent entre les 2 virus sont surlignés ; les 4 acides aminés (positions 121, 165, 172 et 175) impliqués dans le tropisme intestinal de JcDV sont surlignés en rouge [30]. B) Gros plan de la capside de JcDV montrant la topographie au niveau de l’axe de symétrie 5 et la localisation des 8 acides aminés de VP4, exposés à la surface, qui diffèrent de ceux de GmDV (en rouge) (modifié à partir d’une image fournie gracieusement par A. Bruemmer [10]). Les chiffres « 5 » et « 2 » en blanc sur la figure indiquent, respectivement, la position de l’axe de symétrie 5 (au centre du plateau en étoile) et celle de l’axe de symétrie 2. Barre d’échelle : 2 nm.
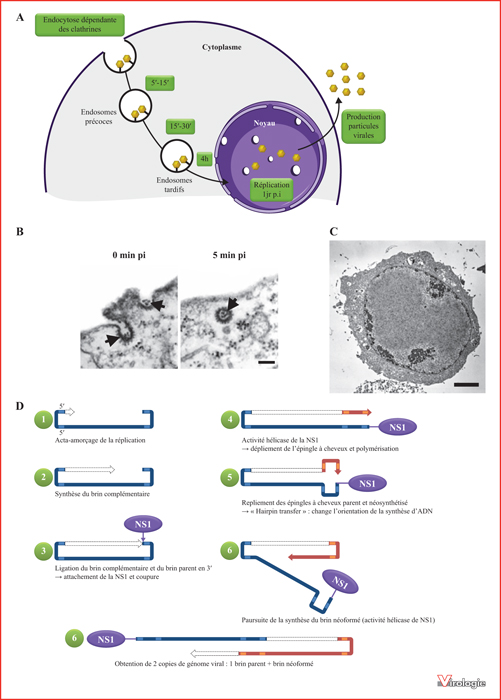
Cycle de multiplication de JcDV et stratégie de réplication des Parvoviridæ. A) Cycle de multiplication de JcDV dans les cellules Ld652Y (d’après [47]). B) Micrographie électronique montrant l’entréepar endocytosedes particules virales dans les cellules Ld652Y. Les flèches indiquent les puits (0 min p.i.) et les vésicules (5 min p.i.) recouvertes de clathrine. Barre d’échelle : 100 nm. [47]. C) Micrographie électronique montrant la densonucléose caractéristique de l’infection densovirale dans les cellules Ld652Y à 6 jours p.i. Barre d’échelle : 10 μm. (Marc Ravallec). D) Schéma présentant la stratégie de réplication des Parvoviridæ.Le brin parental est en bleu, le brin néosynthétisé en rouge. (Adapté de [15].).
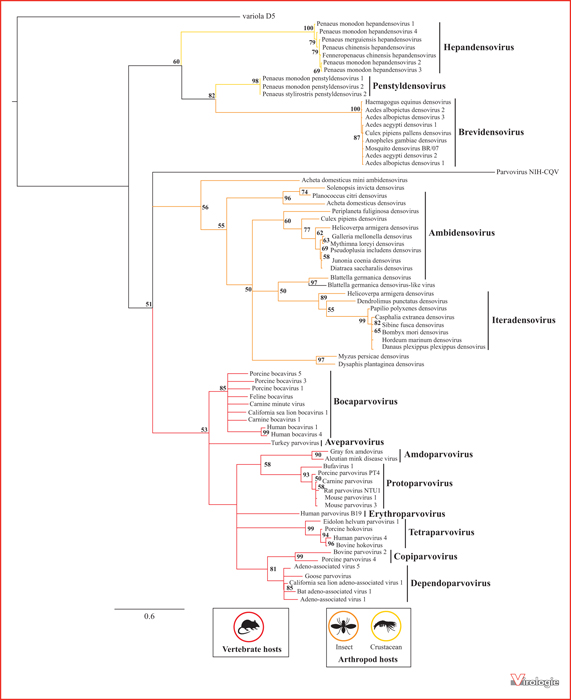
Arbre phylogénétique des Parvoviridæ. Cet arbre regroupe les densovirus répertoriés à ce jour par l’ICTV, ainsi que des parvovirus représentatifs de chacun des genres des Parvovirinæ. Il a été construit par la méthode du maximum de vraisemblance en utilisant PHYML 3.1 à partir d’un alignement de 127 acides aminés appartenant au domaine hélicase SF3 de la protéine NS1. La robustesse des nœuds de l’arbre a été testée par ré-échantillonnage selon la méthode du bootstrap (avec 500 itérations). La protéine D5 du virus de la variole humaine est utilisée comme groupe externe. Seules les valeurs de bootstrap supérieures à 50 % sont indiquées. L’échelle représente le taux de substitution par acide aminé et par position. Les genres auxquels appartiennent les virus sont indiqués à droite. Les hôtes associés sont représentés par des couleurs de branches différentes (vertébrés en rouge, insectes en orange et crustacés en jaune). Les branches terminales notées en noir correspondent à des Parvoviridæ dont les hôtes sont inconnus.
1 Université de Montpellier,
UMR 1333 DGIMI « Diversité,
Génomes et Interactions Microorganismes-Insectes »,
place Eugène-Bataillon,
34095 Montpellier cedex 5, France
Les densovirus (DV) sont des parvovirus d’arthropodes responsables d’épizooties chez les insectes et les crustacés. Structurellement simples, ces petits virus à ADN présentent une grande diversité de séquences et d’organisations génomiques, diversité probablement sous-estimée au regard des récentes découvertes de ces virus dans des hôtes inattendus. Les densovirus représentent un modèle de choix pour étudier à différentes échelles les interactions virus-hôtes et leurs évolutions. Nous proposons de revisiter les connaissances fondamentales sur les densovirus qui ont essentiellement été établies par des approches mécanistiques et envisageons les nouvelles perspectives d’études permises par des approches plus globales. Pour conclure, nous décrivons les applications possibles de ces virus comme outils biologiques, notamment pour le contrôle de populations d’insectes dits « nuisibles ».