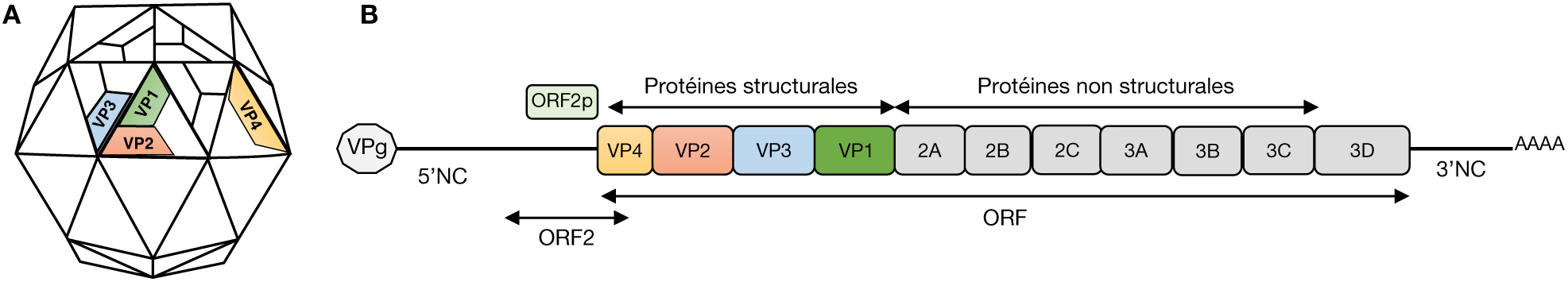
Représentation schématique du génome et de la capside des entérovirus. Les entérovirus (EV) sont des petits virus (25 à 30 nm de diamètre) non enveloppés appartenant à la famille des Picornaviridae. A) La capside de symétrie icosaédrique est formée d’un arrangement de 60 capsomères comprenant chacun quatre protéines structurales (VP1, VP2, VP3 et VP4). Selon les données de diffraction des rayons X réalisée sur des particules virales congelées à -196 °C, la protéine VP4 est localisée à l’intérieur de la capside. B) Le génome des EV est un ARN simple brin positif (7400 bases environ) comportant un grand cadre de lecture (ORF) flanqué d’une région non codante (NC) à chaque extrémité. Un cadre de lecture plus court (ORF2) a été récemment identifié en amont de l’ORF. La région 5′-NC est liée de façon covalente à une protéine appelée VPg de 22 à 24 acides aminés et la région 3’-NC est terminée par une queue poly(A). L’ORF code pour une polyprotéine qui donne après clivage les 4 protéines structurales VP1-VP4 de la capside et sept protéines non structurales impliquées dans la réplication. L’ORF2 est traduit en une seule protéine appelée ORF2p qui est impliquée dans l’infection des cellules intestinales. NB : les termes ORF2/uORF et ORF2p/UP pour désigner le gène et la protéine respectivement sont encore en concurrence.
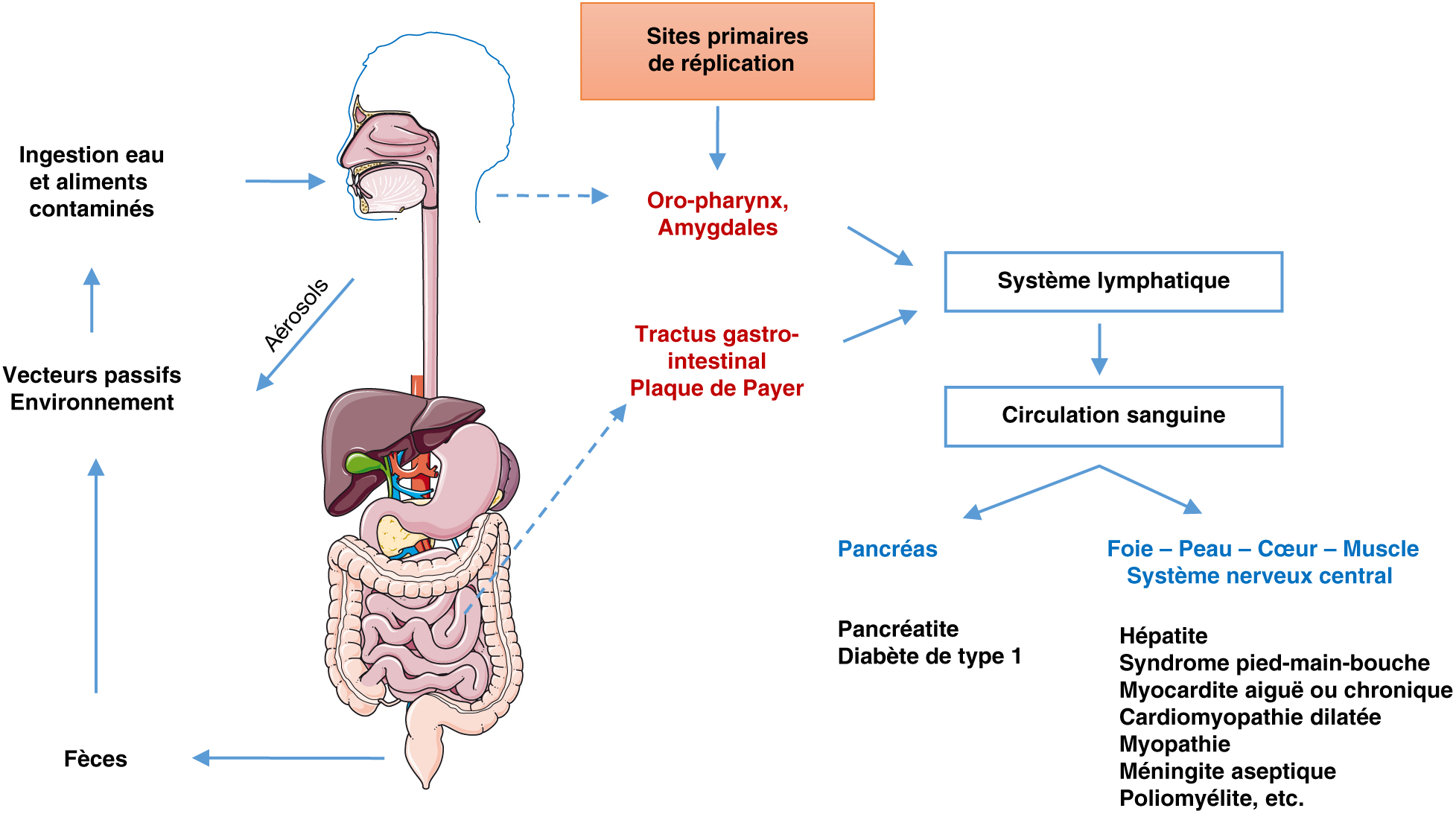
Entérovirus : Transmission, dissémination et pathologies associées. Les entérovirus se transmettent principalement par la voie féco-orale. Une transmission respiratoire ou verticale peut également se produire. Ils se répliquent dans la muqueuse intestinale ou les voies respiratoires supérieures et atteignent des organes cibles par le système lymphatique et la circulation sanguine. Les infections à entérovirus sont généralement asymptomatiques, mais elles peuvent être responsables de symptômes bénins ou de pathologies aiguës et chroniques plus sévères.
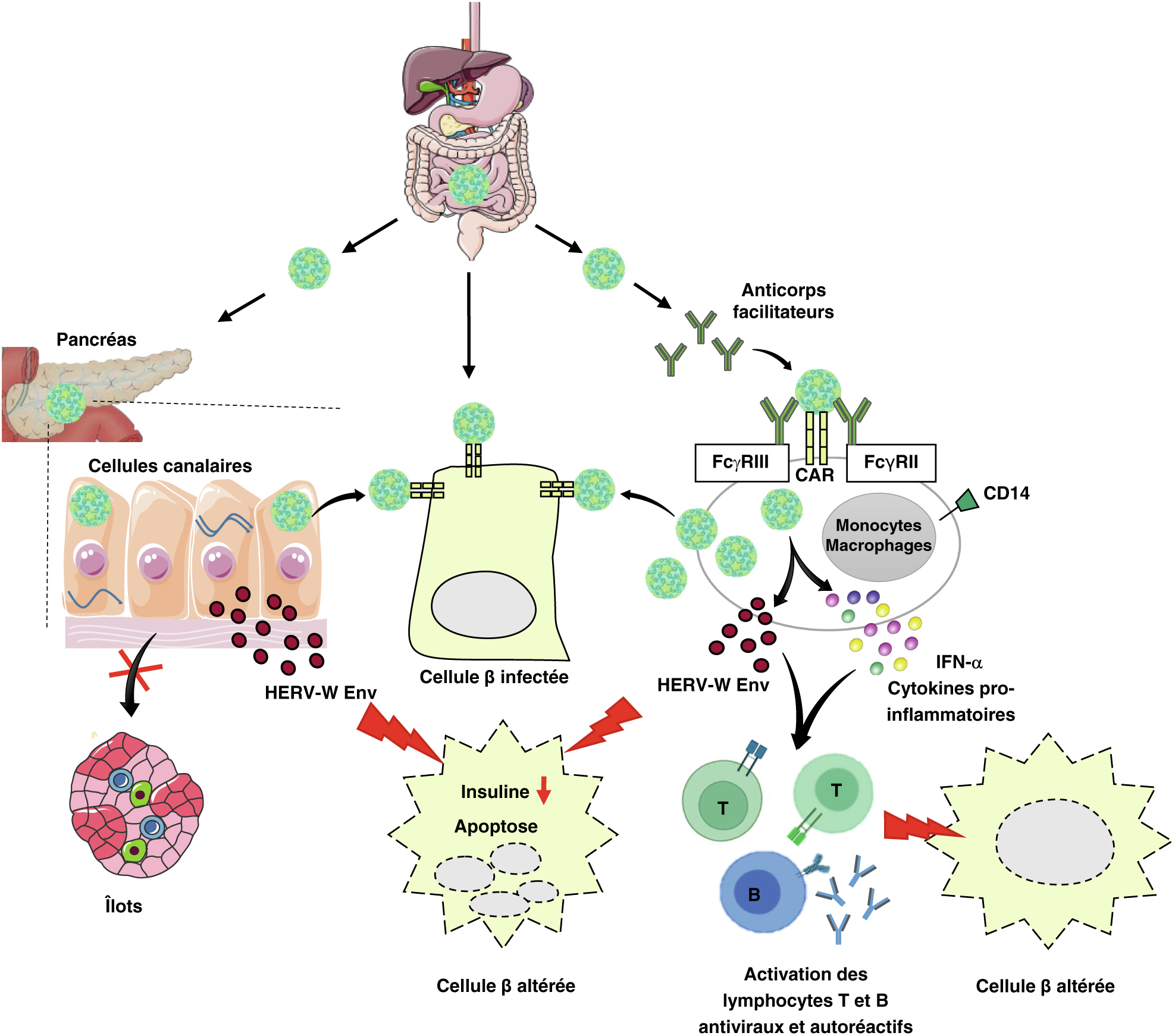
Pathogenèse virale du diabète de type 1 : infection à coxsackievirus B des intestins, du thymus et du pancréas. Après une réplication primaire dans l’épithélium gastro-intestinal et/ou respiratoire, les coxsackievirus B (CVB) atteignent le pancréas et le thymus par le système lymphatique et la circulation sanguine. Les CVB peuvent infecter les cellules β et persister sous forme d’ARN viral double brin qui active un récepteur intra-cytoplasmique, MAD5. Il en résulte une régulation positive de l’expression d’IFNs de type 1, notamment d’IFN-α, capable d’induire une expression accrue des antigènes HLA de classe 1 à la surface des cellules β infectées et des cellules voisines dont la conséquence est la présentation des autoantigènes des cellules β et des antigènes viraux. L’IFN-α ainsi que les cytokines pro-inflammatoires produites provoquent également une inhibition de PCSK2 (enzyme impliquée dans la maturation de la pro-insuline en insuline), une altération de la sécrétion d’insuline, une apoptose des cellules β et une activation des lymphocytes T et B auto-réactifs et antiviraux. L’infection du thymus et en particulier des cellules épithéliales thymiques par le CVB4 peut perturber la tolérance vis-à-vis des cellules β et conduire à la production de lymphocytes T autoréactifs. Les infections par les CVB, répétées et/ou persistantes de la muqueuse intestinale peut favoriser la production et l’activation des lymphocytes antiviraux et autoréactifs. Le mimétisme moléculaire et l’activation de lymphocytes autoréactifs préexistants avoisinants (bystander activation) sont des mécanismes possiblement mis en jeu pour provoquer des altérations des cellules β et un diabète de type 1.
Pathogenèse virale du diabète de type 1 : infection à coxsackievirus B dépendante d’anticorps, rétrovirus humains endogènes et pancréas. L’infection des monocytes et des macrophages par les coxsackievirus B (CVB) peut être facilitée par des anticorps ce qui entraîne la production d’IFN-α et de cytokines pro-inflammatoires capables d’activer des lymphocytes autoréactifs. Les monocytes et les macrophages infectés par le CVB4 peuvent participer à la propagation du virus aux cellules pancréatiques. De plus, le CVB4 active dans les macrophages et les cellules canalaires pancréatiques l’expression d’HERV-W Env qui a des effets délétères sur les cellules β. Le CVB4 persiste dans les cellules canalaires pancréatiques et peut se propager aux cellules β à partir de ces sites. L’infection par le CVB4 des cellules canalaires pancréatiques, altère leur différenciation en cellules productrices d’insuline.
Des études épidémiologiques et expérimentales suggèrent que les entérovirus (EV) et particulièrement les coxsackievirus B (CVB) sont susceptibles de déclencher ou d’accélérer l’apparition de l’auto-immunité anti-îlots et le développement du diabète de type 1 (DT1) chez des individus génétiquement prédisposés. Plusieurs mécanismes non exclusifs mutuellement ont été proposés pour expliquer l’implication des CVB dans la pathogenèse du DT1. Les CVB peuvent infecter et persister dans l’intestin, les cellules thymiques, les monocytes/macrophages, les cellules ductales et β pancréatiques, ce qui entraîne des altérations structurelles ou fonctionnelles de ces cellules. Une réponse inflammatoire chronique et une perturbation de la tolérance vis-à-vis des cellules β dues aux infections à CVB sont capables de favoriser le recrutement et l’activation des cellules T auto-réactives préexistantes et la destruction des cellules β. Plusieurs stratégies vaccinales ou thérapeutiques de lutte contre les infections à EV ont été développées et ouvrent des perspectives pour la prévention ou le traitement du DT1.