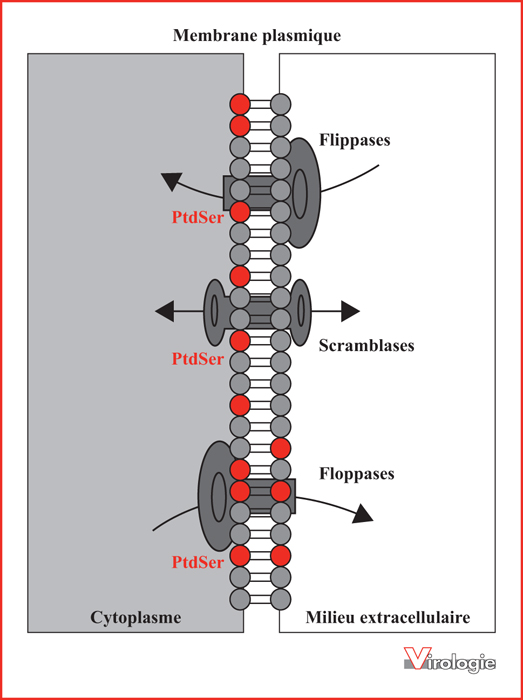
Contrôle de la répartition de la phosphatidylsérine (PtdSer) dans la membrane plasmique cellulaire par les translocases.
La PtdSer est enrichie dans le feuillet interne de la membrane plasmique. L’asymétrie de sa distribution est maintenue sous l’action dépendante de l’ATP de translocases telles que les flippases. Les floppases exercent une action inverse, tandis que les scramblases transportent les phospholipides de façon bidirectionnelle. Ces translocases jouent un rôle important dans l’exposition de la PtdSer lors des étapes précoces de l’apoptose [1, 3].
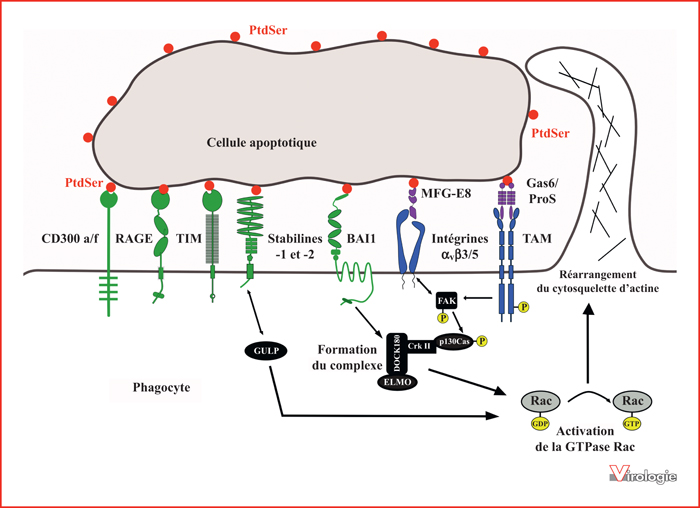
Deux modes de reconnaissance de la phosphatidylsérine (PtdSer) par les récepteurs impliqués dans l’internalisation des cellules apoptotiques.
La PtdSer exposée à la surface des cellules apoptotiques est reconnue directement par les récepteurs des familles TIM, CD300, les récepteurs RAGE, BAI1 et les Stabilines-1 et -2 (en vert). La présence de ligands Gas6 ou ProS et MFG-E8 (en violet) est indispensable lors de la reconnaissance indirecte de la PtdSer par les récepteurs TAM et par les intégrines αvβ3/5 respectivement (en bleu). Certains récepteurs peuvent alors engager différentes voies de signalisation aboutissant à l’activation de la GTPase Rac, impliquée dans les réarrangements du cytosquelette d’actine nécessaires à l’ingestion par phagocytose des corps apoptotiques.
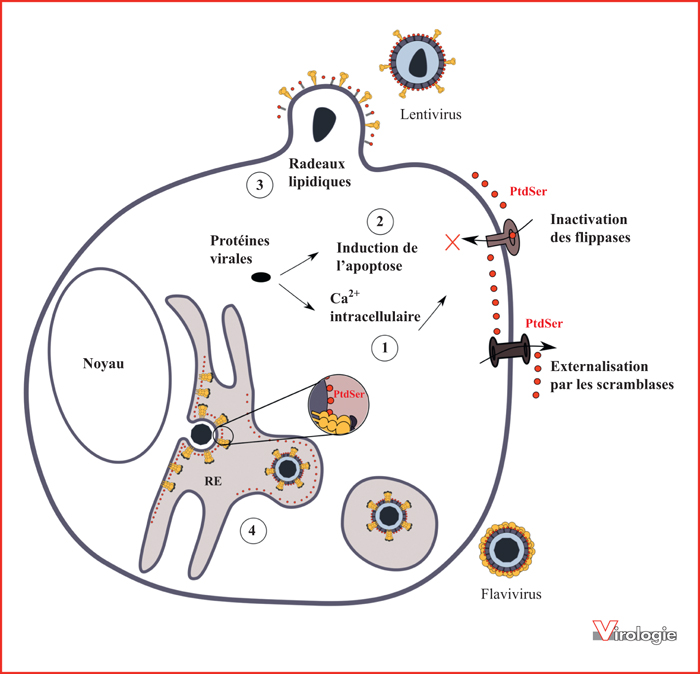
Hypothèses concernant l’acquisition de la phosphatidylsérine (PtdSer) dans les enveloppes virales.
Différentes hypothèses peuvent expliquer l’incorporation de la PtdSer dans les enveloppes virales. Les enveloppes des virus sont dérivées des membranes cellulaires acquises durant l’étape de bourgeonnement et potentiellement enrichies en PtdSer. Par exemple, l’exposition de la PtdSer à la surface des cellules infectées peut résulter soit de la réplication virale pouvant entraîner une augmentation du Ca2+ intracellulaire (1), soit de l’induction de l’apoptose par le virus (2) ayant pour conséquence l’externalisation de la PtdSer par les scramblases et l’inactivation des flippases. Selon les virus, le bourgeonnement peut se produire soit au niveau de radeaux lipidiques à la membrane plasmique [53] dont le feuillet externe peut être transitoirement enrichi en PtdSer (3) soit au niveau du réticulum endoplasmique (RE) naturellement riche en PtdSer (4).
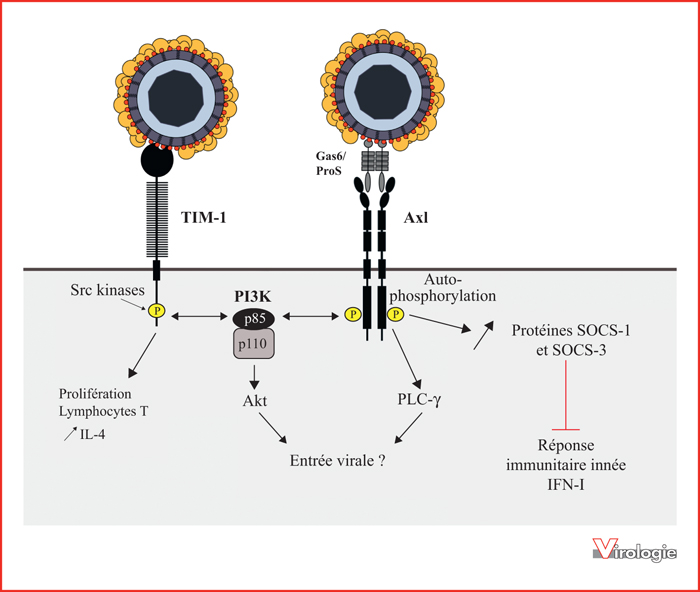
Modèle proposé des voies de signalisation dépendantes de TIM-1 et Axl activées par les virus.
L’interaction directe ou indirecte de la phosphatidylsérine (PtdSer) associée au virus avec les récepteurs TIM-1 et Axl peut permettre au virus d’exploiter des voies de signalisation activées par ces récepteurs et favorisant l’infection. Parmi ces voies, celle impliquant la PI3K, ou encore la PLC-γ peuvent être détournées par le virus afin de promouvoir son entrée. Dans le contexte d’une liaison indirecte, le complexe virus/ligand peut agir comme un supergoniste du récepteur TAM et favoriser son homodimérisation ainsi que son activation par autophosphorylation. Ainsi, l’activation du récepteur Axl induit une augmentation de l’expression des protéines SOCS-1 et -3 impliquées dans l’inhibition de la réponse immunitaire innée, favorisant la réplication virale [50].
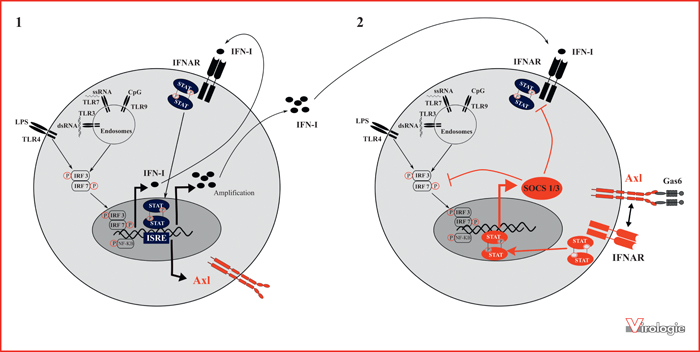
Rôle du récepteur Axl dans l’inhibition de la réponse immunitaire innée.
Inspiré du modèle proposé par Rothlin et al. en 2007 [10]. (1) L’activation des TLRs par un pathogène induit la production et la sécrétion d’interféron de type I (IFN-I). La fixation de 1IFN-I à son récepteur IFNAR conduit à la phosphorylation et l’activation des facteurs transcriptionnels STAT, dont la translocation dans le nucleus induit la transcription des gènes régulés par l’élément de réponse à l’interféron (ISRE), dont le gène codant le récepteur Axl. (2) L’activation d’Axl par son ligand Gas6 induit son association avec IFNAR, permettant ainsi de détourner la voie IFNAR/STAT et stimuler la transcription des gènes SOCS. Les protéines SOCS-1 et -3 sont des inhibiteurs des voies de signalisation dépendantes des TLRs, terminant ainsi une boucle de rétrocontrôle négatif de la réponse immunitaire innée IFN-I.
L’élimination des cellules apoptotiques, ou efferocytose, est un processus cellulaire essentiel et très conservé qui repose principalement sur la reconnaissance de la phosphatidylsérine (PtdSer) exposée à la surface des corps apoptotiques par le phagocyte. Depuis une dizaine d’années, plusieurs études ont montré que de nombreux virus mimaient ce processus biologique majeur pour infecter leurs cellules cibles en exposant la PtdSer à la surface de leur enveloppe lipidique. Ce concept de « mimétisme apoptotique » a été renforcé récemment car de nombreuses familles de virus exploitent des récepteurs liant directement ou indirectement la PtdSer pour s’attacher aux cellules cibles et initier leur cycle infectieux. Cette revue fait le point sur les avancées majeures dans ce domaine de recherche et discute l’implication des récepteurs à la PtdSer, notamment les familles TIM (T-Cell Immunoglobulin and Mucin domain) et TAM (Tyro3, Axl, Mer), dans le processus d’entrée des virus.