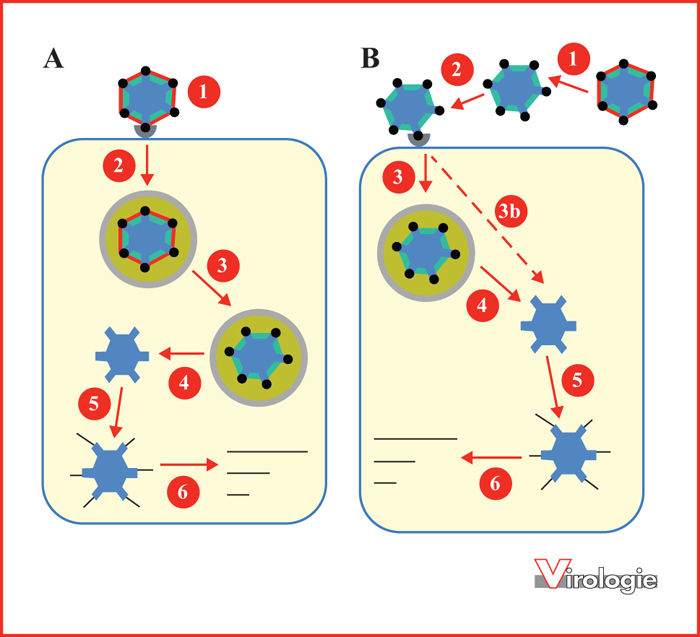
Fixation et entrée de réovirus. A) Le schéma présente les principales étapes de l’entrée du virus à partir du virion. Dans un premier temps, (1) celui-ci se fixe à la surface cellulaire par la protéine trimérique de spicule σ1 (sphère noire). Après l’endocytose (2), l’activité des protéases lysosomales permet le retrait de la protéine σ3 (en rouge) et le clivage de la protéine μ1 (en vert) (3), permettant à la particule de traverser la membrane de l’endosome. La sortie de l’endosome (4) s’accompagne de l’élimination de la protéine σ1 et de la portion résiduelle de la protéine μ1. Ces dernières étapes impliquent la participation de protéines cellulaires jouant le rôle de chaperon moléculaire. La particule virale se retrouve finalement dans le cytoplasme sous forme de nucléoïde (ou core), particule complètement dépourvue de protéines de la capside externe et transcriptionnellement active (5). Le nucléoïde ainsi relargué dans le cytoplasme permet la synthèse des trois classes d’ARN messager viral (6). B) Le schéma représente l’entrée virale lorsque le virion est tout d’abord partiellement décapsidé par l’action de protéases extracellulaires. (1) La particule sous-virale infectieuse (ISVP) se fixe à la membrane (2) pour soit être internalisée par endocytose (3) ou, alternativement, pénétrer directement au travers de la membrane plasmique (3b), contournant dans les deux cas le besoin en protéases endosomale/lysosomale. Dans tous les cas, le nucléoïde est par la suite généré (4) pour permettre la transcription comme dans le cas du virion (5, 6).
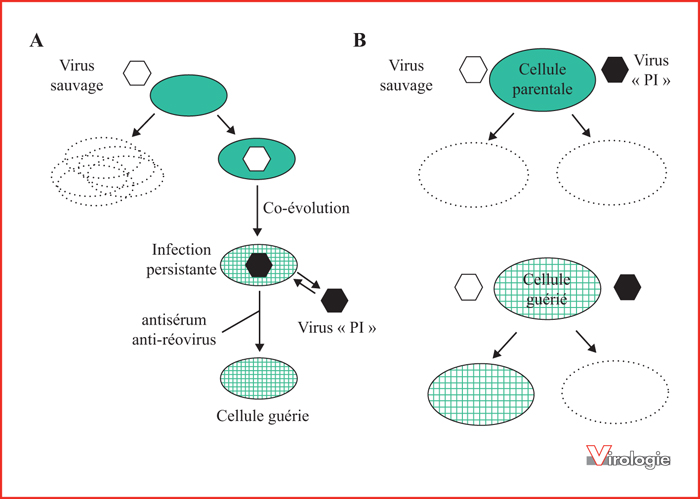
Établissement de la persistance virale et coévolution virus-cellule. A) Lors de l’infection de cellules par un réovirus de type sauvage, la majorité des cellules seront tuées et détruites par cytolyse (en pointillés). Toutefois, quelques cellules peuvent être épargnées et devenir infectées de manière persistante (en vert). Avec le temps, la cellule pourra évoluer (quadrillée) tout comme le virus pour devenir un virus dit PI (persistent infection) (en noir). Il s’établit un état d’équilibre avec libération de particules virales et réinfection constante des cellules. De telles cellules peuvent être « guéries » de leur infection par ajout d’un antisérum neutralisant qui prévient la réinfection des cellules. B) Les cellules originales parentales peuvent être infectées et détruites avec le virus sauvage ou le virus récupéré de cellules infectées de manière persistante (virus PI). Par contre, les cellules dites « guéries », ayant été en contact avec le virus pendant une longue période de temps lors de la persistance virale, sont résistantes au virus sauvage de départ, mais sensibles au virus PI.
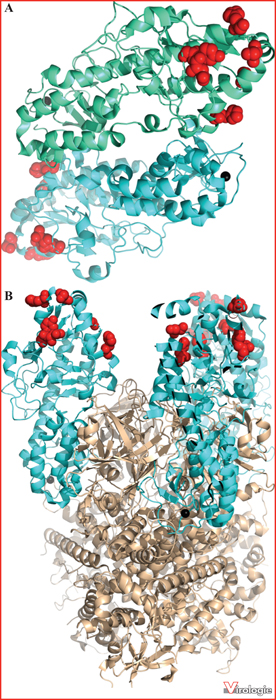
Position des substitutions d’acides aminés de la protéine σ3. A) La structure cristallographique de la forme dimérique de la protéine σ3 (en vert et cyan) (PDB : 1FN9) est présentée [19]. B) L’hétérohexamère composé de trois molécules de la protéine σ3 (cyan) à la surface du virion, coiffant trois molécules de la protéine μ1 (en brun pâle) (PDB : 1JMU) est présenté [20]. Les acides aminés substitués chez divers virus PI sont indiqués sous forme de sphères rouges sur la structure de la protéine σ3.
Position des substitutions d’acides aminés de la protéine σ1. La structure cristallographique de la forme trimérique de la protéine σ1 (PDB : 6GAP) [42] est présentée. Les acides aminés substitués chez divers virus PI sont indiqués sous forme de sphères rouges, la substitution démontrée comme importante chez le virus PI isolé de cellules Vero (VeroAV) est indiquée. Les 26 premiers acides aminés et les 215 derniers acides aminés, comprenant la structure globulaire de la protéine, sont absents de cette structure (lignes pointillées). La flèche bleue indique la position de la fin de la forme tronquée observée chez le virus PI isolé de cellules de fibrosarcome humain HT1080. La position de la région de fixation aux acides sialiques [43] est aussi indiquée (trait vert).
Représentation schématique des étapes présumées d’établissement et de maintien de la persistance. A) Dans les cellules L929, des changements dans la région externe de la protéine σ3 (en rouge) au sein de l’hétérohexamère de σ3-μ1 semblent essentiels à l’établissement de la persistance virale (1). Des changements subséquents au niveau de la région hélicoïdale (en rouge) de la protéine de fixation σ1 apparaîtraient par la suite (2). B) Dans les cellules Vero, les premiers changements apparaissent au niveau de la région de fixation aux acides sialiques sur la protéine σ1 (en rouge) (1) et sont suivis d’autres changements (en rouge) au niveau de σ1 et de μ1 au sein de l’hétérohexamère σ3-μ1 (2). Plus de détails sont présentés dans le texte.
Université de Montréal,
Département de microbiologie, infectiologie et immunologie,
Université de Montréal
CP 6128, Succ. Centre-villeMontréal, QC, Canada H3C 3J7
Bien qu’étant peu pathogène chez l’humain, le réovirus de mammifère a été, et demeure, un modèle privilégié d’étude de la réplication et de la pathogenèse virale. Généralement considéré comme étant cytolytique, ce virus peut parfois établir des infections persistantes à long terme en culture cellulaire. Dans ce contexte, il constitue en fait un des modèles parmi les plus utilisés pour démontrer la coévolution entre virus et cellules. Bien qu’initialement surtout limitées au modèle des cellules fibroblastiques de souris L929, les dernières années ont vu apparaître d’autres études au sein de différents types cellulaires. L’utilisation de la persistance virale pourrait devenir un outil de prédilection pour isoler de nouveaux virus mieux adaptés à des utilisations virothérapeutiques, par exemple en tant qu’agents oncolytiques contre des cancers humains ou animaux. Une meilleure compréhension du phénomène de persistance, spécialement des gènes viraux impliqués, s’avère donc essentielle. Le développement de nouveaux outils, tel que l’utilisation de la génétique inverse, apparaît très prometteur pour l’atteinte de ces objectifs. Cette dernière approche permet en effet d’établir la signification biologique de mutations retrouvées chez les virus sélectionnés lors de la persistance virale.



 Cette œuvre est mise à disposition selon les termes de la
Licence Creative Commons Attribution - Pas d'Utilisation Commerciale - Pas de Modification 4.0 International
Cette œuvre est mise à disposition selon les termes de la
Licence Creative Commons Attribution - Pas d'Utilisation Commerciale - Pas de Modification 4.0 International