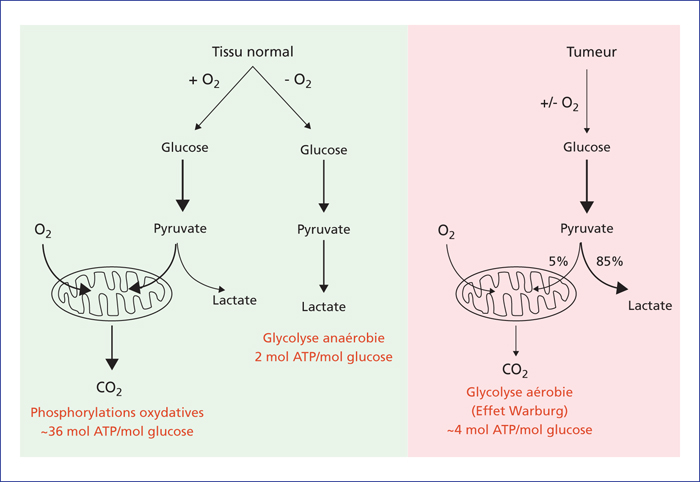
Représentation schématique des différences métaboliques entre les cellules normales et les cellules cancéreuses (schéma adapté de la revue de Vander Heiden et al.[5]). Représentation schématique des phosphorylations oxydatives, de la glycolyse anaérobie et de la glycolyse aérobie connue sous le nom d’« effet Warburg ». Dans le cas des tissus normaux (non prolifératifs), le glucose est métabolisé en pyruvate via la glycolyse et en présence d’O2, et le pyruvate est complètement oxydé en CO2 dans les mitochondries pour produire de l’ATP. En revanche, en absence d’O2, le pyruvate est transformé en lactate, ce qui permet à la glycolyse de continuer, mais en produisant peu d’ATP. Dans le cas des tumeurs, le pyruvate produit lors de la glycolyse est transformé en lactate, et ce, indépendamment de la présence ou de l’absence d’O2, même si les mitochondries sont toujours fonctionnelles.
Diagram of the metabolic differences between normal and cancer cells (adapted from Vander Heiden et al.[5]). Oxidative phosphorylation and anaerobic and aerobic glycolysis is depicted, demonstrating the “Warburg effect”. In the case of normal (non-proliferative) tissues, glucose is metabolised to pyruvate through glycolysis, and in the presence of O2, pyruvate is completely oxidised to CO2 in the mitochondria to produce ATP. In contrast, in the absence of O2, pyruvate is transformed into lactate, which enables glycolysis to continue, but produces little ATP. In the case of tumours, during glycolysis, the pyruvate produced is transformed into lactate and this occurs independently of the presence or absence of O2, even if mitochondria remain functional.
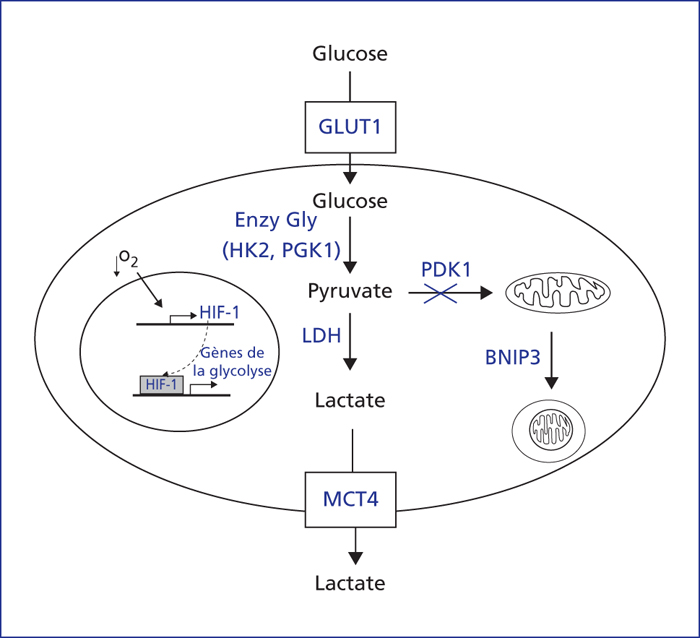
Le régulateur majeur de la reprogrammation métabolique des cancers est HIF-1. HIF-1 [18] induit l’expression de GLUT1 pour faciliter l’entrée de glucose, ainsi que les enzymes de la glycolyse HK2 et PGK1 pour augmenter la dégradation de glucose en pyruvate, la LDH pour produire le lactate et MCT4 pour évacuer ce lactate. En outre, HIF-1 induit l’expression de BNIP3 pour augmenter la mitophagie, et celle de PDK1 pour inhiber la pyruvate déshydrogénase, afin de réduire l’utilisation de pyruvate par les mitochondries.
The major regulator of metabolic reprogramming of cancers is HIF-1. HIF-1 [18] induces expression of GLUT1 to facilitate the entry of glucose, as well as glycolytic enzymes, HK2 and PGK1, which increases the degradation of glycose to pyruvate, LDH to produce lactate, and MCT4 to evacuate this lactate. Moreover, HIF-1 induces the expression of BNIP3 to increase mitophagia and PDK1 to inhibit pyruvate dehydrogenase, in order to reduce the use of pyruvate in mitochondria.
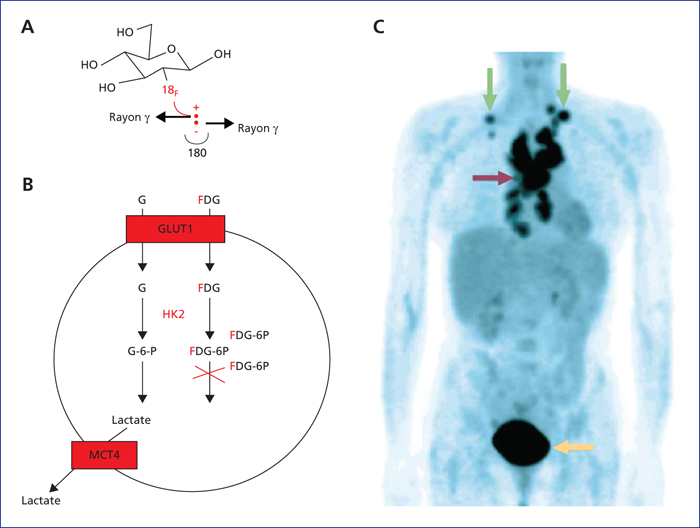
Utilisation de la glycolyse en tomographie par émission de positrons. (A) Structure chimique du 2-(18F)-fluoro-2-désoxy-D-glucose (FDG). Le radioélément 18F émet des positrons (β+) qui produisent de l’énergie sous forme de deux rayons gamma à 180° l’un de l’autre. (B) Le FDG est transporté dans les cellules comme le glucose ; en revanche, après phosphorylation par les HK2, le FDG bloque la progression de la glycolyse, induisant son accumulation dans la cellule qui sera à l’origine du contraste sur l’imagerie. (C) Image en tomographie par émission de positrons après injection de FDG chez un patient atteint d’un lymphome. Les tumeurs médiastinales (flèche rouge) et sus-claviculaires (flèches vertes) montrent une forte accumulation de FDG. La vessie montre également un fort signal car le FDG est éliminé par le système urinaire entre 30 et 60 min après injection (flèche jaune).
Utilisation of glycolysis in positron emission tomography. (A) The chemical structure of 2 (18F)-fluoro-2- deoxy-D-glucose (FDG). The radio-element 18F emits positrons (β+</sup) which produce energy in the form of two gamma rays at 180̊ from one another. (B) FDG is transported into cells like glucose, however, after phosphorylation by HK2, FDG blocks the progression of glycolysis, leading to its build-up in the cell which creates the contrast on imaging. (C) Positron emission tomography after injection of FDG in a patient with a lymphoma. The mediastinal tumours (red arrow) and sub-clavicular tumours (green arrows) show a strong build-up of FDG. The bladder also produces a high signal because the FDG is eliminated through the urinary tract between 30 and 60 minutes after injection (yellow arrow).
MitoLab, Mitochondrial Medicine Research Centre, UMR CNRS 6015, INSERM U1083, Institut MitoVasc, Université d’Angers, CHU Bât IRIS/IBS
Rue des Capucins
49933 Angers cedex 9
France
Au cœur des nombreuses adaptations des cellules cancéreuses à leur environnement se trouve la reprogrammation métabolique. En effet, les tissus cancéreux utilisent principalement la glycolyse pour produire de l’énergie, mécanisme 18 fois moins efficace que celui de la phosphorylation oxydative utilisée par les tissus « normaux ». Cette reprogrammation métabolique est mise en place pour soutenir les divisions anarchiques et ininterrompues des cellules cancéreuses. Un certain nombre de gènes orchestrent ce processus et présentent une expression anormale et souvent corrélée avec l’agressivité tumorale. Après un rappel sur les fonctions des mitochondries, cette revue présentera les différences métaboliques majeures entre tissus sains et cancéreux, ainsi que les nouvelles approches diagnostiques et thérapeutiques les exploitant au mieux. Considérer le cancer comme une entité avec son métabolisme propre pourrait nous aider à développer de nouvelles stratégies efficaces en association avec des thérapies existantes.

 Cette œuvre est mise à disposition selon les termes de la
Licence Creative Commons Attribution - Pas d'Utilisation Commerciale - Pas de Modification 4.0 International
Cette œuvre est mise à disposition selon les termes de la
Licence Creative Commons Attribution - Pas d'Utilisation Commerciale - Pas de Modification 4.0 International